![]()
Семейството на кучетата Canidae, обединява видове като вълка, чакала, лисицата и кучето. От деца сме израснали с приказките за ненаситния вълк, хитрата лисица и вярното куче. Какво би станало, ако освен кучето и лисицата бе станала верен другар на човека? В днешно време това е напълно вазможно да се разбере, тъй като има вече домашни лисици и те по нищо не отстъпват на кучетата.
Лисиците, които общо наброяват 37 вида, като цяло са отделени таксономично от кучетата в отделен таксон Vulpini. Лисиците са били и остават неизменна част от културата на човека. Няма митология и фолклор, който да не включва лисицата като символно животно. От древни времена хората са ловили лисици, най-вече заради кожите. Лова е останал като спорт и до днес, а кожи все още се търгуват успешно на световния пазар, въпреки дейността на природозащитниците. Разпространението на лисиците по света е вървяло успоредно със хората. В по-нови времена, лисицата е интродуцирана в Австралия, което е създало проблем в екеологичното равновесие.
Същинските лисици са обединени в род Vulpes . Дванадесетте вида, макар да имат доста сходни белези, не малко имат и различия. Доста видове, понеже обитават трудно достъпни места, не са проучени добре, както по отношение на екологията, така и физиология, етология и генетика. Не е изяснен и филогенетичния статус, като и филогеографията на повечето от тях. Таксономично, при всеки един от видовете, е необходимо да се изяснявани статуса на подвидовете. Все още има много насоки за научни и природозащитни разработки на тези така добре познати ни животни.
![]()
Пустинна лисица, Фенек, fennec fox ,Vulpes zerda, Zimmerman, 1780
![]()
Най-малката лисица в света се нарича фенек, Vulpes zerda. Тя живее в Северна Африка, Сахар, Арабския полуостров и северно Мароко, далеч от селища и градове. Тъй като се е адаптирала за живот в пустинята е наричана още пустинна лисица. Името и на арабски означава „лисица”, докато zerda произлиза от гръцката дума сух – xeros, поради сухите условия при които живее. Той е много мъничък - с дължина - 37 до 43 см и височина - 18 до 21 см, той тежи само 1-1.5 кг. Фенека има големи уши, до 15 см дълги, които му служат да се охлажда, има красива пясъчно оцветена козина, жълтеникава на гърба и по-бяла на корема. Сетивните и органи са много чувствителни, а тялото и е много пъргаво и чевръсто и така се справят с предизвикателствата на живот на открито.
![]()
Очите са големи с тъмни ириси и специална ретина, даваща много добра зрителна острота дори през нощта. Добре са развити обонянието и слуха. Муцуната е малка, къса, леко издължена, горната устна носи няколко двойки черни мустаци. Челюстите са въоръжени с остри зъби, особено кучешките. Опашката е 18-31 см дълга, пухеста , черна към върха. Ако трябва да го сравним с другите лисици, бихме могли да кажем, че това животно е джудже, но много пъргаво и силно, успешно успяващо да лови малки животни, живеещи в пустинята. Плячката му се състои от различни животни, живеещи в Сахара – бозайници, насекоми, влечуги, мишки, плъхове, гекони, птички, яйца, членестоноги, скакалци. Фенека ловува на залез слънце, тъй като е нощно животно. Има специфичен начин на лов, понеже живее далеч от водни източници, той преследва внимателно плячката, скача и я захапва за врата, после я донася до бърлогата си, където необезпокоявано я изяжда. Към менюто му се включват и различни плодове, корени и грудки, от които си набавя необходимите течности. Фенека прави леговището си от коренища, треви и малко храсти, които може да намери в околностите. Издава различни звуци, с които комуникира със събратята си. Обикновено живее на семейни групи от по 10 индивида, като маркира територията си , за да я предпази от натрапници. Мъжкия защитава малките си на територията и може да стане много агресивен срещу други животни. Пустинните лисица са моногамни животни, които се размножават веднъж годишно. Женската се разгонва само за два дни и показва това свое състояние на мъжкия. След чифтосването, бременноста трае около 50 дни. Раждат се от 2 – 3 слепи и безпомощни малки. През първите 5 – 6 седмици, малките бозаят, а мъжкия не се допуска до бърлогата. Новородените тежат около 50 г. Малките отварят очи и започват да се окосмяват на около 2 седмици след раждането. След две три седмици ушите им започват да растат, като отначало са много малки, но после стават големи, защото растат по-бързо от тялото. На 1 – 2 месечна възраст майката ги отбива, на 3 месечна започват сами да си търсят храна на дълги разстояния, а на 9 – 11 месечна възраст достигат полова зрялост и могат да се размножават.
Тази лисица е най-красивото животно в пустинята, което лесно се опитомява, лесно се хваща и продава заради козината си или като домашно животно. В плен е наблюдавано да живее до 13 – 14 годишна възраст. Местните жители го наричат: „Душата на пустинята”.
![]()
Антоан дьо Сент Егзюпери пише в писмо до сестра си Диди през 1918г от Кейп Джуби, за фенек, който отглежда и обожава. Той също се натъква на пустинната лисица, когато неговия самолет пада в пустинята в Сахара през 1935г. Именно тези му срещи с това странно животно го вдъхновяват за образа на лисицата в „Малкия принц”.
Фенека Баха е описан в детската книжка на Елинор Хофман „Mischief in Fez”, публикувана през 1943г. . Според тази история, фенека е описан според арабските и мавританските традиции,че се отглежда в дома, за да пази от джинове и други демонични духове. Фенека е символа на екологията на Тунис. Фигурата на това животно, облечено в синьо - бял костюм може де се види в почти всеки град на тази страна. Фенека е изобразен на монета от ¼ динара. В следните художествени произведения на известни писатели се срещат интересни сведения за фенека:
Майн Рид. Млади пътешественици или Момчета-ловци на Север (The Young Voyageurs: or The Boy Hunters in the North) (1854) (глави 11, 13).
Антоан дьо Сент-Экзюпери. Terre des hommes, Земя на хората (1939) (глава VII).
Антоан дьо Сент-Экзюпери. «Малкия принц».
Анри Троая. «Гладът на лъвчетата», «Крушение».
Сергей Лукьяненко. Прозрачните витражи
Firefox for mobile — логотип и кодово име Firefox for mobile.
Феликс Кривин. Лисицата фенек.
Франк Хърбърт - "Дюн"– от малкото животни, обитаващи Аракис
Видео:
https://www.youtube.com/watch?v=4AhkhPCXc3A
Южноафриканска лисица, Капска лисица, Cape fox, Silver fox, Vulpes chama, Smith 1833
Този вид е широко разпространен в централните и западни части на Южна Африка. Обитава откритите тревисти пространства, с разпръснати ниски храсти , гористи региони, както и полупустинни и пустинни. Характерна за Зимбабве и Ботсвана. През последните деситилетия, ареала и се разширява югозападно и достига крайбрежието на Атлантическия и Индийския океан. Описана е за Ангола, Ботсвана, Лесото, Намибия и Южна Африка.
Малка лисица със стройно тяло и пухкава опашка с черен връх. Мъжките са около 5% по-големи от женските. Цялостната окраска е сребристо сива, като долните крайници, главата, гърба на дългите уши са червеникаво кафяви до бледо кафеникави. Има бели косми по бузите и върховете на ушите. Дължината на тялото достига 45 -61 cm, опашката е с дължина 30 - 40 cm, теглото 3.6 - 5 kg.
Южноафриканската лисица е нощно животно, като е най - активна на сумрак, преди залез или на разсъмване. През деня се укрива в подземни дупки, цепнатини или гъсти храсталаци. Тя може да изкопае дупка сама, въпреки че предпочита да използва вече изкопана от друг вид. Живеят поединично, като формират двойки през брачния период. Не са строго териториални, но маркират обитавания от тях район. При опасност защитават района около бърлогата си, особено ако има малки. По принцип не са шумни, но комуникират със меки звуци, виене, пронизителни звуци и други. Когато алармира издава вой завършваш с кратко излайване, а когато се чувства нападната ръмжи и плюе в нападателя си . За да покаже, че е развълнувана, вдига нагоре опашката си, обикновено начина на положение на опашката издава степента на вълнение.
![]()
Тази лисица е всеядна, като менюто и включва растения и животни. Въпреки, че предпочитат безгръбначни и малки бозайници като гризачи, те ловуват и влечуги, зайци, паяци, птички. Хранят се и с яйца, ларви на бръмбъри,бръмбъри, скакалци, мърша, насекоми и плодове.
![]()
Брачния период при южноафриканската лисица в някой региони не е строго сезонна, докато в други е сезонна. Основния сезон е пролетно-летния. След бременност от 51 - 53 дни, се раждат малките през август-октомври, най-често септември. Обикновено се раждат от 1 до 6 малки лисичета, в изкопаните или разширени дупки – бърлоги. При раждането си тежат едва 50 – 100 гр. Лисичетата се хранят от двамата родители, но основно мъжкия подсигурява храната. При опасност и двамата родители защитават своите малки. Малките се включват в лова на възраст около 16 седмици, а напускат семейството на около шест месечна възраст. Обикновено семейството включва възрастните и малките, но различните семейства се смесват при наличия на храна. Може да се застъпят няколко поколения, но по-често женската гони порасналите лисичета, когато очаква новото поколение. Южноафриканските лисици достигат полова зрялост на около 9 месечна възраст. Продължителност на живота средно 6 години, но има наблюдения и до 10 годишна възраст.
![]()
Смята се, че южноафриканската лисица регулира числеността на гризачите. Основни нейни врагове са големи грабливи птици, каракал, хиена, лъв и леопард. Често се подават на болести като бяс и гана, а в по-ново време стават жертви на капани за проблемни животни. Не малко загиват по пътищата от автомобили. На някой места са гонени и преследвани като вредители, тъй като често се бъркат с чакала. Рядко се продават техни кожи и почти не могат да се видят в магазини за кожи, основно защото са малки животни. В Ботсвана се използват заедно с други кожи за направата на традиционните одеала (kaross), но в последно време тази направа е силно намаляла, поради индустриалното производство. Средно годишно се счита, че загиват около 2500 индивида, но въпреки това в червената книга на IUCN е описан като слабо засегнат вид.
Видео:
https://www.youtube.com/watch?v=pdP6xxDYvfE
Тибетска лисица, Тибетска пясъчна лисица, Tibetan sand fox, Vulpes ferrilata, Hodgson, 1842
![]()
Ендемичен вид, разпространен в степите и полупустините на Тибетското плато. Също така се среща в Непал, на север от Хималаите, в северозападна Индия, китайските провинции, граничещи с Тибетската автономна област. Тези лисици са наблюдавани в планините до 5200 м надморска височина. Тибетските лисици не се срещат там, където отсъстват черномуцунести сеносъбирачи (Ochotona curzoniae), които са основната им храна.
![]()
Една от най-малките лисици. Зъбите са добре развити, с необичайно големи кучешки зъби, по-големи от на всички останали лисици. Тялото на тибетската лисица е покрито с дебела, мека козина с плътен пухест подслой, който добре ги защитава от силните ветрове. Муцуната изглежда донякъде квадратна, заради гъстата козина около шията. Цвета по страните се изменя от рижаво - кафяво на върха до сиво отдолу, което създава илюзията на ивици по страните на животното. Гърлото, коремната област, ивица от гърлото до корема и върха на опашката е бяла на цвят. Дължина на тялото на възрастна тибетска лисица от 60 до 70 см. Опашката е с дължина от 30 до 45 см. На тегло са особи от 4 до 5,5 кг. В кариотипа си имат 36 хромозоми.
Тибетските лисици живеят в изкопани бърлоги или дупки под скали или скални насипи, цепнатини и водят доста потаен начин на живот, за който е много малко известно. Тези животни ловуват по двойки /мъжки и женски/ и делят плячката с своя партньор. Имат тънък слух, позволяващ отдалече да намират любимата си храна черномуцунести сеносъбирачи. Териториалноста при тези лисици е изразена слабо и за разлика от други, те не защитават своя участък от другите лисици. Наблюдавано е, че много двойки живеят по съседсво и ловуват в общи участъци. До 2006 няма направени видоематериали на тази лисица, поради труднодостъпните региони, където живее и от гледна точка опазване на вида. Екип от BBC успява да снима и включи материал за тибетската лисица в документалния филма Планета Земя , в серия 7 - Великите равнини.
Както казахме, любимата храна на тибетските лисици са черномуцунести сеносъбирачи (Ochotona curzoniae), които съставляват до 80% от менюто им, но освен това те могат да се хранят и с гризачи, зайци, малки птички, гнездящи по земята и техни яйца. Каквото успеят да уловят като плячка, подходяща за храна, например насекоми, влечуги. Разнообразяват с плодове, например плодчета от Ephedra. Не се отказват и от мърша.
Сезона на размножаване настъпва през февруари. Двойките се образуват в края на първата година от живота и остават за цял живот. Живеят и възпитават децата си заедно. Бременноста е с продължителност 50-60 дни, средно 55 дни. След раждането женската не напуска бърлогата поне няколко седмици. В бърлогата се раждат от 2 до 5 малки, слепи и голи лисичета, всяко тежи около 60-120 гр., това обикновено става през май. Младите не се появяват навън в течение на няколко седмици след раждането, докато не пораснат. Точната продължителност на кърменето не е уточнена. Счита се, че продължителността на живота на тези лисици е от 8 до 10 години, но в естествените местообитания средната прод ължителност е около 5 години.
Кожата на тибетската лисица се използва от жителите на някой области на Тибет за правене на шапки, които надежно могат да защитават от вятър и дъжд, но нямат голяма комерсиална стойност. Тибетските лисици имат съществена роля в регулация на числеността на гризачите и други малки животни. Обсъжда се също вероятността да проветряват почвата, ровейки своите леговища.
Макар в червената книга на IUCN ,видът да е описан като слабо застрашен, популацията на тези лисици не е многобройна и е възможно да намалее драстично в бъдеще. Опасност за нея представляват действията на правителството на КНР , което спонсорира кампанията по отравянето на любимата храна на тези лисици черномуцунести сеносъбирачи на територията на Тибетското плато. Още една опасност за тибетската лисица са домашните кучета на коренното население на Тибет. В близкото минало доста лисици са избити от ловци, но сега това е ограничено.
![]()
Видео:
https://www.youtube.com/watch?v=lUKe6_dS4IE
http://en.wikipedia.org/wiki/Planet_Earth_(TV_series)
http://ru.wikipedia.org/wiki/%D0%9F%D0%BB%D0%B0%D0%BD%D0%B5%D1%82%D0%B0_%D0%97%D0%B5%D0%BC%D0%BB%D1%8F_(%D1%84%D0%B8%D0%BB%D1%8C%D0%BC)
В серия 7 на филма на ББС – Планета Земя
Американската прерийна лисица, swift fox, Vulpes velox, Say 1823
![]()
Американска лисица или американски корсак, прерийна лисица, Vulpes velox. Името и на латински velox и на английски swift, означава бързонога.
Американската лисица е малка сива, с големи уши и широка, къса муцуна. Височина около 30 см, дължина на тялото 37—53 см, дължина на опашката 22—35 см, тегло 2—3 кг. Мъжките са малко по-големи от женските. Козината е гъста, недълга, опашката е пухкава. Обичайно козината е оцветена в светло сиво, с рижи отенъци по страните и краката. Лятото козината става по-рижава. На гърдите и гърлото, козината е по-светла. Крайчето на опашката е черно. На муцуната и от двете страни има черни петна.
Първоначално американската лисица се е срещала в откритите тревисти равнини и в пустините на юго-западна Канада, от Великите равнини до щата Тексас. На територията на Канада тя напълно изчезва през 1930 г. В сегашно време големи популации на тази лисица са се съхранили в САЩ от изток от Скалистите планини – Уайоминг, Колорадо, Канзас и Ню Мексико, също така в Монтана, Северна и Южна дакота, Небраска, Оклахома и на северозапад в Тексас. По-рядко в Айдахо и Орегон. На изток в Ню Мексико и на запад в Тексас има зона на хибридизация между Vulpes velox и Vulpes macrotis.
![]()
Американската лисица живее в откритите нискотревисти прерии, в тревистите равнини и в сухите полупустини. Както всички обитатели на аридните области, лятото води нощен или сумрачен начин на живот, а зимата може да излиза на слънце и денем. Лятната дневна жега или ветровитите дни тя преживява в дълбоки подземни дупки, които изкопава сама, обикновено по склоновете на хълмовете. Рядко заемат изоставени дупки на язовци.
Доскоро се смяташе, че американските лисици не са териториални, нямат граници на участъците, които обитават. По нови изследвания показват териториалност, като един участък заема площ от 1,5 до 5 км² и в тези места, където са малки, обикновено участъците се припокриват. Американските лисици се хранят основно с гризачи и зайци, също така птички, които гнездят на земято, влечуги и плодове. Зимата важен източник на храна е мършата, оставаща от по-големи хищници. Лятото ядат много насекоми, главно бръмбари и скакалци / до 50% от менято/. Лисиците сами стават плячка на койоти.
Американските лисици водят общо взето потаен начин на живот, за който се знае малко. Много са боязливи и в случай на опасност стремително бягат, развивайки скорост до 60 км/ч, като лесно менят посоката на движение, оттук и наименованието им бързоноги лисици.
Описан е вокален репертоар, като са идентифицирани осем различни вокализации : териториален, агонистичен, покорно хленчене, покорно бърборене, прекопулативен, социален, предупреждаващ, издаващ вълнение. Звуците са различни, някой от тях наподобяват лаене, подлайване.
Те са моногамни животни, двойките се формират в октомври - ноември. Сезона на размножаване продължава до декември / на юг в САЩ/ и до март / Канада/. Малките се раждат, след 50 - 60 дневна бременност от март до май. Обикновено са от 3 – 6 на брой, слепи, голи и безпомощни, като правило се раждат повече мъжки, но въпреки това като цяло в популацията се поддържа равновесие между броя на мъжките и женските. Проглеждат на 10 – 15 ден, в дупката остават до края на първия месец от живота си . Бозаят до 6 – 7 седмица. На тримесечна възраст могат да ловуват наравно с родителите си . Мъжкия помага в отглеждането на потомството. Семейството се разпада само през септември - октомври, а младите започват да търсят свои участъци. Половата зрялост достигат до година за мъжките, а женските до две години. Продължителността на живота в природата - 3 - 4 години, в плен – до 13 г.
![]()
Този вид е описан в червената книга на IUCN като слабо засегнат вид, но въпреки това числеността на вида и неговия ареал, чувствително е намалял през XX век. В XIX век – началото на XX век, американските лисици активно са ловени, заради кожата, макар тяхната кожа да не се е ценяла високо, поради грубостта си и малкия си размер. Често тези лисици попадат в капани, поставени за обикновените лисици или койоти. Роля в съкращаването на числеността на американските лисици е имало също така използването на отровни вещества за изтребване на гризачи и разрушаване на характерната им среда на местообитание – нискотревната прерия.
В началото на XX век, американските лисици бързо изчезват от територията на Канада, последния екземпляр е убит в провинция Саскачеван през 1928 г. През 1978 г. В Канада този вид е обявен за измрял. От 1984 г. Действащи програми по вторично заселване на тези лисици, водят до това, че сега на територията на Алберта и Саскачеван живеят около 350 индивида. Статуса на популацията на американската лисица на територията на САЩ силно варира, в зависимост от подвида и ареала.
Описани са следните подвидове:
Други автори описват значителна географска вариация, която съществува между американските лисици на север (V. velox hebes) и на юг (V. v. velox).
Всички гореописани подвидове са таксономично непотвърдени и подлежат на изясняване.
Американската лисица фенотипично и екологично е сходна с лисицата джудже, kit fox (Vulpes macrotis), като между тях има хибридна зона в западен Тексас и източно Ню мексико. Някой морфометрични сравнителни анализи, както и протеин - електрофоретични данни водят до извода, че тези лисици принадлежат към един вид и редица автори ги обединяват в един вид Vulpes velox. Противно на това, други мултивариативни морфометрични сравнения, както и анализ на мтДНК секвенции, подкрепят становището за отделни видове. Двата вида лисици са доста близки до полярната лисица, Arctic fox (Alopex lagopus), и тази генетична асоциация е с най- близки родствени връзки, спрямо останалите лисици, въпреки това тези видове остават систематично отделени. Тази филогенетична близост, може да се обясни с това, че най – вероятно видообразуването е протекло твърде скоро, за да може да се определи съществена генетична разлика. Липсват по-нови изследвания в насока определяне на генетично родство , по специално между американската и лисица и лисицата джудже, в бъдещ план едно обширно изследване на последователностти на мтДНК, а също и по нуклеарни локуси, биха съществено допринесли за разрешаването на таксономичния статус и по-категорично определяне на взаимовръзките между трите вида лисици.
Лисица джудже, kit fox, Vulpes macrotis Merriam, 1888
![]()
Лисицата джудже е най-малката американска лисица. Има очевидно големи уши, къса козина , с жълтеникаво до сиво оцветена глава, гръб и страни. Плешките и външната страна на крайниците са с кафеникаво – жълтеникаво оцветяване, а корема, гърдите и вътрешната страна на крайниците са бели до жълтеникави на цвят. Края на опашката е черен. По врата, крайниците и корема може да имат пухести просветлявания.
Описани са осем подвида:
— V. m. arsipus - югоизточна Калифорния, южна Аризона и южна Сонора, Пустинната лисица джудже, понеже се среща в пустинята Мохаве
— V. m. Devia – южна Долна Калифорния
— V. m. macrotis - югозападна Калифорния - Южнокалифорнийската лисица джудже изчезнал подвид от 1903г
— V. m. mutica - Долината Сан Хоакин в Калифорния, Санхоакинската лисица джудже
— V. m. neomexicana - Ню Мексико, западен Тексас и северозападна Чихуахуа
— V. m. nevadensis - региона Great Basin в САЩ
— V. m. tenuirostris - северна Долна Калифорния
— V. m. Zinseri – северно централно Мексико
Като цяло систематиката на този вид и систематичния му статус, както упоменахме по горе, подлежи на ревизия.
Днес лисицата джудже обитава пустинните и полупустинните региони на западна Северна Америка. В САЩ от южна Калифорния до западно Колорадо и западен Тексас, на север до южен Орегон и Айдахо. В Мексико през полуостров Долна Калифорния и северна Сонора и Чихуахуа до западен Нуево Леон, на юг в северен Закатекас. Предпочитат сухите места, тревисти с халофилни почви, срещат се от 400- 1900 м надморска височина, въпреки че избягват терени с възвишения. За изкопаване на бърлоги предпочитат рохкави почви. Макар в по- малка степен обитават и градски условия, агрикултурни райони, предимно градини и зеленчукови насаждения.
![]()
Тези лисици се хранят предимно с гризачи, зайцевидни и насекоми. Основната им плячка включва кенгурови плъхове от род Dipodomys spp., прерийни кучета от род Cynomys spp., калифорнийски зайци Lepus californicus и американски зайци Sylvilagus spp. Допълнително менюто включва птички, влечуги и мърша. По-рядко се хранят с растителна храна, понякога с кактусови плодчета.
Джуджевидните лисици са адптирани към живот в сухи и горещи условия. За да отдели излишната топлина, тялото им е с по-голяма повърхност , спрямо масата и големи уши. Така има възможност да се отдели топлина, без да се губи вода. Предимно имат нощна активност, а през деня са в бърлогата си. Набавят си необходимата вода от тяхната храна.
Предимно са моногамни животни, рядко се наблюдава полигамия. Двойките обикновено са за цял живот. Младите лисици от предишното котило, обикновено женски, може да не напуснат семейството и да останат в помощ на отглеждането на новото котило. Лисиците джудже не са строго териториални и районите им често се застъпват, въпреки че основната част се използва само и единствено от едно семейството. Териториите им се простират от 2.5km² до 11.6km².
Тези лисици лаят, когато видят наближаващ хищник или викат малките си, понякога издават предупредително ръмжене, срещу натрапниците навлизащи в тяхната територия. Лисици джудже в плен са наблюдавани да издават звуци при затворена уста, когато са разтревожени. Мирисната маркировка не е все още изучавана.
![]()
Брачния им период трае от декември до януари. От средата на февруари до средата на март се раждат малките лисичета, които са обикновено от 1 до 7 , средно 4 в едно котило. Фертилитета зависи годишно от наличието на храна за всяка от възрастите. След около 4 седмици малките се показват извън бърлогата, бозаят до около осмата си седмица, а на 3 – 4 месечна възраст вече се хранят с родителите си. На 8 месечна възраст вече могат да напуснат семейството и да водят самостоятелен живот.
Лисиците джудже използват леговището си целогодишно и имат по няколко леговища на своя територия. Въпреки, че сами изкопават дупки за леговища, понякога могат да заемат и променят дупки на други животни, като на прерийни кучета, катерици, кенгуру плъхове и язовци. Рядко могат да имат леговища и в структури, построени от човека, като тръби, но малките при всички случаи се раждат в дупки, направени в земята.
Основни врагове на лисиците джудже са койотите, по-рядко са нападани от грабливи птици, язовци, червен рис, обикновенната лисица, диви кучета. В Мексико могат да бъдат застрелвани по грешка, но не са преследвани. В САЩ голям брой са избити повреме на различни програми за контрол на хищниците, главно койоти и вълци. Кожата им няма висока стойност и рядко са преследвани заради нея. В САЩ са регистрирани 1200 индивида, избити заради кожата през 1994-1995 г. В Мексико тези лисици са продавани като домашни любимци нелегално.
В червената листа на IUCN са регистрани като слабо засегнат вид. Въпреки това лисиците джудже са с намаляващи популации. Санхоакинската лисица джудже, V. m. mutica е приета за застрашен в САЩ, тъй като нейните хабитати са доста разпръснати и намаляват за сметка на обработваемите земи. Лисицата джудже се счита за засегната в някой щати, като Колорадо и Юта, където има програми за защита на този вид. Този вид се счита за застрашен в Калифорния и Орегон. В Мексико се счита за уязвим, където се смята ,че около 40% от популацията е намаляла, поради разширение на обработваемите земи.
Като цяло са необходими допълнителни изследвания за този вид, по отношение на биогеграфия, екология, етология, физиология, генетика и филогеография.
Лисица на Рюпел, Пясъчна лисица, Rüppell's fox, Vulpes rueppellii,Schinz 1825
![]()
Първоначално е била класифицирана в род Canis, като Canis rueppellii, но в последствие преместена в рода на истинските лисици. Името rueppellii идва от латинското Rueppellius – от името на германския колектор Едуард Рюпел. Разговорното пясъчна лисица, често се използва и за лисицата корсак и тибетската пясъчна лисица, затова не е за предпочитане.
Разпространена в биотопите на пустинните и полу - пустинните региони на Северна Африка / на север от 17°N/ от Мароко и Мавритания до Египет и Сомалия, на север се ограничава от северните граници на пустиня Сахара. Разпространена е и на Арабския полуостров, на изток до Пакистан (68°E) и на северозапад до Израел и Йордания. Типичния им хабитат са пясъчните и каменисти пустини. В Суадитска Арабия се срещат на открити и каменисти хабитати с рядка тревиста растителност.
![]()
Лисицата на Рюпел е дълга 40-52 см, приблизително тегло 1,7 кг. Тя е една от най-малките лисици, с по-слабо телосложение от обикновенната червена лисица V. Vulpes и има по слаби крака. Ушите са дълги и големи, участват в отделянето на топлината на тялото, донякъде приличат на тези на фенека, но им липсват тъмните петна откъм гърба им. Цвета на козината варира. Главата е бежова до бледо пясъчна на цвят. Ушите и муцуната са бледи, с черни снопчета мустаци. Цвета на гърба варира от бледо пясъчен до сив и дори червеникав. Страните и долната част на тялото е бледо, краката са бежови до светлобежови, а лапите са покрити изцяло с косми, вероятно за по - лесно придвижване върху пясъка. Черните пръски по гърба, завършват с черна черта по дължина на опашката. Самата опашка завършва с бял край, което се счита за диагностичен белег. През зимата козината е по-тъмна, докато през лятото е светла.
Описани са следните подвидове:
— V. r. Rueppellii – в Египет и Судан
— V. r. caesia - в северозападна Африка и Западна Сахара
— V. r. cyrenaica - включва V. r. cufrana? – в Либия, югозападен Египет, северозападен Судан
— V. r. Somaliae – в Еритрея и Сомалия
— V. r. sabaea - в Средния Изток и Арабския полуостров
— V. r. zarudnyi - в Балучистан
Диетата им включва безграбначни, гризачи, гущери, змии, птички и плодове. Малките бозайници са основна част от менюто и ако намалеят или изчезнат, ги заменят с насекоми от род Coleoptera.
Способността им да оцеляват при живот в хипер – аридни условия , където възможността да пиеш вода е изключително рядка, се основава на различни екологични, етологически и физиологически приспособления.
Малко се знае за начина им на живот, но като цяло е наблюдавано, че лисицата на Рюпел живее на групи от по 3 – 15 индивида, което най-вероятно е семейство от няколко поколения. В Оман е наблюдавано, че теригорията на тези лисици се простира до 69km². Средно обаче територията е от 10,2 до 16.3km².
Двойките са моногамни. Брачния период е от декември до февруари. Бременността трае 51-53 дни, след което се раждат 2 – 3 слепи и безпомощни малки. Малките бозаят до 6-8 седмична възраст. Те растат под земята, за да бъдат предпазени от хищници. Полова зрялост достигат до 9-10 месечна възраст. Като цяло екологията и етологията на вида не са добре проучени.
![]()
Фрагментацията, деградацията и загубата на хабитати, директното и индиректното преследване от ловци, използването на отрови, са основната причина за намаляването на популацията на лисицата на Рюпел в Мароко. В Израел този вид е на път за изчезне, поради конкуренцията на червената лисица, която разширява своя регион, следвайки човека в пустинята Негев.
Рядко, но все пак, видът е убиван за храна или заради кожата. Кожи са намирани на пазарите за кожи. В повечето държави лисицата на Рюпел присъства в защитени райони. С изключение на Суадитска Арабия, където не е нелегален лова и отстрела, както и отравянето на хищници, в повечето от останалите държави, където се среща този вид, има ограничения. В червената листа на IUCN видът е включен като слабо засегнат.
Бледа лисица, Африка́нская лиси́ца, Pale fox ,Vulpes pallida, (Cretzschmar, 1826)
![]()
Първоначално видът е бил описан като Canis pallidus Cretzschmar, 1827. Пустинните лисици, фенека(V. zerda)., лисицата на Рюпел (V. rueppellii) и бледата лисица Vulpes pallida са били приемани като една обща група пустинни лисици и дори обединявани с индийската лисица (V. bengalensis) и южноафриканската (V. chama).
Бледата лисица е разпространена в полупустинния район Сахел на Африка, граничещ със Сахара, от Мавритания и Сенегал, през Нигерия, Камерун и Чад до Червено море. Южната граница на географския ареал стига до северна Гвинейска лесосавана.
![]()
Малка, бледо оцветена лисица, със сравнително дълги крака и големи уши. Бледа, удължена муцуна с относително дълги мустаци и черен пръстен около очите. Големи уши, бели отвътре и червеникаво- кафяви отвън. Тялото е кремаво бяло до пясъчно бежово, относително късокосместо, с черна или червеникаво кафява линия на гърба. Страните са по-бледо оцветени, отколкото гърба, като са пухкаво бели от вътрешната страна и червеникави крака. Имат дълга пухкава опашка, червеникаво кафява с черен връх и тъмно петно над опашната жлеза. Тялото на бледата лисица е дълго 38-45 см, дължината на опашката е 28-29 см. , а на тегло е — 1,5-3,6 кг.
Описани са няколко подвида, по долу са дадени четири от тях:
— V. p. pallida - в Судан, от Кордофан до Донгола
— V. p. edwarsi - в Мали, Сенегал
— V. p. harterti - северна нигерия, на север до Аир на Нигер и на запад до Буркиниа Фасо
Обитават пустинните и полупустинните региони, но достигат и по-влажните райони на Гвинейската лесосавана, което предполага, че вида е по-екологично пластичен. Често се наблюдава и близо до населени места, където е по-вероятно да си намери храна.
Добре развитите му молари, предполагат, че тази лисица се храни предимно с растителна храна, плодове. Също така в менюто и влизат различни гризачи, птички, гнездящи по земята, малки влечуги и безгръбначни.
Адаптирани са за живот в сухи условия, като си набавят необходимата вода от плодове и храната като цяло. Въпреки това не се продължително издръжливи на сухи условия.
Наблюдавани са да живеят на групи, вероятно семейството с малките. Мъжкия и женската дълбаят голяма дупка за бърлога, където ще отглеждат малките си. Дълбока 2 – 3 м , с вътрешна камера, обикновено в сухи песъчливи места. В плен е наблюдавана бременност от 51-53 дни, след която се раждат от 3 – 6 малки, които се кърмят от 6- 8 седмици.
Бледата лисица е най-малко изследваната лисица и много все още има да се научи за нея, по отношение на екология, етология, физиология, генетика, филогеография и т.н.
В червената книга на IUCN видът е включен като слабо засегнат.
![]()
Бенгалска лисица, Индийска лисица, Bengal fox, Indian fox, Vulpes bengalensis, Shaw, 1800
![]()
Индийската лисица е ендемична за Индийския субконтинент. Ареала и се простира от възвишенията на Хималаите в Непал до южния край на полуостров Индустан. В северната част на полустрова, ареала се простира от провинция Синдх в Пакистан до северен Бенгал в Индия. Индийската лисица предпочита полусухи, проветриви простори, с оскъдна растителност, където лесно ловят плячка и копаят леговища. Избягват гъстите гори, терени с висока растителност и същински пустини.
![]()
Индийската лисица е средна по ръст. Дължина на тялото 50 см, дължина на опашката 25-30 см., тегло 3-4 кг.Ушите са пропорционално големи, с тъмно кафяво оцветяване откъм гърба. Носа и устните са черни, а очите имат тъмни пръски. Муцуната е оцветена в тъмно към черно откъм горната си част и към очите. Окраската е сива, варираща от жълтеникавосива до сребристосива, без ръждиво червени косми, както е окраската на червената лисица. Гръбно е по-тъмно оцветена, докато долната част е бледо кремаво до мръсно бяло. През зимата палтото става по пухкаво и гъсто. Краката са сравнително слаби и леко рижави .Опашката е повече от половината на дължината на тялото. Върха на опашката е черен. Лисицата държи опашката си нормално при бавно движение, хоризонтално при бяг и почти вертикално при внезапна промяна на движението.
Индийската лисица е всеядна и може да се храни с почти всичко, което успее да улови. Менюто включва основно насекоми – щурци, крилати термити, скакалци, мравки, бръмбъри и паяци; малки гризачи, птички и техните яйца, гущери, змии и зайци. Освен животинска, лисицита се храни и с вегетарианска храна, като плодове, корени и грудки.
Тези лисици водят сумрачен или нощен начин на живот, като обикновено изчакват да се свечери, за да излязат на лов. В дни на умерена температура или валежи, могат да излязат на лов и през деня. Въпреки, че живеят по двойки, обикновено ловуват и се хранят поединично. Основната социална единица е семейната двойка, която се формира с цел създаване на потомство и се задържа за няколко години. Около нея могат да останат подрастващите малки и така да се оформи цяло семейство.
Индийските лисици издават различни звуци, за защита на територията си, за аларма и комуникация помежду си . Маркират територията си с мирисни вещества.
Брачния им период е от декември до януари. През този период мъжкия издава непрекъснато интензивни звуци, рано сутрин на зазоряване или късно вечер на смрачаване, също така и през ноща. Бременността е с продължителност от 50-53 дни, раждат в периода от януари до март. Малките са от 2 до 4, слепи и безпомощни, тежащи от 50- 100 гр. И двамата родители участват в отглеждането на малките. Грижата за подрастващите продължвата 4-5 месеца. След като станат достатъчно самостоятелни малките напускат семейството и започват да водят самостоятелен живот. Продължителност на живота: от 6 до 8 години. Бърлогите си лисиците изкопават сами, рядко използват чужди, които разширяват. Самите бърлоги представляват сложен комплекс от дупки и тунели.
![]()
Основния враг на индийските лисици са хората, особено номадските племена Nari kurava и техните кучета. Представителите на тези племена навлизат в териториите на лисиците, успешно имитират техните звуци, като по този начин ги примамват. Убиват ги заради месото, зъбите, ноктите и кожата. Често използват саморъчно направени и обмазани с животинска мазнина бомби, използват пушек, кучета и мрежи или блокират отворите на дупките с камъни. Племената waghri и koli убиват индийските лисици, за ядене.
Не е известно масово използване на тези лисици с комерсиална цел, макар да се намирани на индийските пазари кожи, нокти и зъби, които се продават за медицински цели и за правенето на магии.
Загубата на хабитати и болестите също сериозно се отразяват на популацията на тези лисици. В червената листа на IUCN е включена като слабо засегнат вид.
Видео:
https://www.youtube.com/watch?v=LRs9E_8CvYg
Лисица Корсак , corsac fox , Vulpes corsac, Linnaeus, 1768
![]()
Корсак или степна лисица, още пустинна лисица е разпространена в степите, полупустините и отчасти в пустините на Югоизточна Европа и Азия, заедно с обикновенната и афганската лисици. Корсака е разпространен от северен Иран, Казахстан, Афганистан и Средна Азия до Монголия и североизточен Китай.
![]()
Корсака прилича на обикновенната лисица, но е видимо по-малък, с по-големи уши и по-високи крака. Дължина на тялото 50—60 см, дължина на опашката 25—35 см, тегло 4—6 кг. Муцуната е къса, силно заострена, скулите са широки, ушите големи, широки в оснавата си, а към краищата заострени. Обичайната окраска е светла, сива или рижо- сива, срещат се и отенъци на червено, коремно е бяла или жълтеникава, крайчето на опашката е тъмно кафяв или черен, под брадата – светла. Имат изразен сезонен морфизъм в дължината на козината, като през зимата е дълга и пухеста, а лятото – къса и прилепнала. От обикновенната лисица, корсака се отличава по тъмния край на опашката, а от афганската лисица по по-късата опашка и бялата окраска на подбрадата и долната устна.
Описани се следните подвидове:
- V. c. corsac - северната част на ареала до предАлтайската степ, не по далеч на юг от северния край на Аралско море
— V. c. turcmenica - степите на Средна Азия и Казахстан, северен Авганистан и североизточен Иран, предполага се, че се припокрива с V. c. kalmykorum.
— V. c. scorodumovi - Руското Трансбайкалие, Монголия и Китай
- V. c. Kalmykorum – Волго уралските степи и дясната страна на басейна на река Волга
![]()
През кватернера ареала на вида е бил много по-голям. През ранен плейстоцен неговия предшественик V. praecorsac е обитавал територията на Австрия и Унгария. В края на плейстоцена корсака е бил разпространен от Швейцария до северен Китай. От края на плейстоцена - ранен холоцен, ареала се стеснява от запад поради климатични промени.
Филогенетични изследвания, основани на секвенции на цитB гена на мтДНК, показват, че генетичната разлика между тибетската лисица и корсака е значително малка, в сравнение между тибетската и обекновенната, червена лисица. Може да се каже, че тези резултати показват, че тибетската лисица е по-близка с корсака отколкото с обикновенната лисица. Според молекулярния часовник, червената и тибетската лисица дивергират преди около 4,8 – 6,4 млн години, докато корсака и тибетската лисица девергират преди 2,8 млн години. Като се съпоставят тези данни с геологичната история на Тибетското плато, може да се каже, че тибетската лисица и корсака са дивергирали през третата фаза на формирането на Хималаите, когато Тибетското плато е било на 2000 м. надм.вис. След този период настъпва бързо повишаване на Хималаите и респективно Тибетското плато, което е основната причина за разделянето и обособяването на двата вида лисици.
![]()
Корсака предпочита хълмистите местности с невисока растителност, полупустини и сухи степи, зимата малоснежни и с плътна снежна покривка. По-малко се срещат в лесостепната зона и в пустините, навлиза в предпланините. Избягва гъста растителност, гори и високотревисти полета. Площта на индивидуалния участък може да достигне 35-40 км², в протежение на който имат мрежа от дупки, проходи и места на маркиране.
Корсака живее в дупки, което е обусловено от местообитанието му – засушливи райони с горещи дни и хладни нощи през лятото и сурови зими. За жилище използва дупки, които преустройва , на мармоти, язовци или земни катерици, като по-рядко сами копаят нови. Леговището му е плитко и недълбоко, с дължина на 2,5 м и с няколко входове.
Корсака ловува поединично, предимно на сумрак, но по неволя може да прояви и дневна активност. Има много добро обоняние, зрение и слух. Ловува бавно, като дебне плячката, върви срещу вятъра, прокрадва се или я надбягва. Не допуска лесно човек до себе си, обикновено го избягва, крие се, а ако не може се прави на умрял.
Основно се храни с малки гризачи / полевки, мишки, тушканчици, пустинни плъхове/, влечуги, насекоми, птички и техни яйца. По рядко земни катерици, таралежи и зайци. При недостиг на храна, може да се храни и с отпадъци и мърша. Растителна храна почти не яде. Издържа продължително време без вода. През зимата, поради намаленото количество храна и трудностите в намирането и, числеността на популацията може да спадне няколко пъти. В много райони през есента корсаците отиват на юг, понякога след стадата сайги, които отъпкват снега и така ги облекчават в придвижването и лова. Масови миграции на корсаци могат да бъдат предизвикани от степни пожари, катастрофално измиране на гризачи и др. По време на такива миграции, корсаците се появяват далеч извън границите на своя ареал и дори навлизат в градовете.
Основни врагове са вълка и обикновенната лисица. Вълците ловуват корсаци, които въпреки способността си да развиват добра скорост (до 40—50 км/ч), скоро се уморяват и забавят темпа. От друга страна, корсака дояжда остатъците от убити от вълка животни. Обикновената лисица се явява основен хранителен конкурент на корсака.
Лисицата корсак издава звуци като кучетата и лисиците: лаят, вият, джафкат, ръмжат и др., като звуците се използват за различна комуникация помежду им, за аларма и предпазване.
![]()
Корсаците са моногамни животни, двойките остават за цял живот. Брачния период е през януари – февруари, обикновено през нощта и се съпровожда с лаенето на самците и сблъсъци за млади или сами самки. Чифтосването става в бърлогата. Бременността продължава 52-60 дни, а малките се раждат през март - април. Котилото може да бъде от 2 до 11-16 малки, най-често 3-6. Новородените са слепи, покрити със светло кафяви косми и тежат около 60г, дължина на тялото 13-14 см. Проглеждат на 14 – 16 ден, на месец започват да се хранят с месо. На 4 – 5 месечна възраст достигат размерите на възрастни и се разселват. Когато застудее, младите корсаци отново се събират и зимуват в една бърлога. Полова зрялост достигат на възраст 9 - 10 месеца. Продължителност на живот в природни условия е неизвестна, предполага се до 6 години.
Корсака е предмет на търговия с кожи, като главно се използва зимната кожа. Известно е, че в Казахстан се лови от бронзовата ера до сега. Казахите и киргизите са използвали в 13 век кожите на корсака като разменна стока. И сега корсака си остава традиционен ловен обект с грейхаунди / тази/, ловни соколи и скални орли. Допринася за изтребването на гризачите. Точни данни за числеността на популацията липсват. Бракониерството оказва сериозно влияние на тази численост. През 19 век, на година са избивани над 10 000 индивида заради кожата, в някои години по 40 000 – 50 000.
![]()
Видът е включен в червената книга на IUCN като слабо засегнат. Също така е включен и в регионални червени книги в Русия: Башкирия, Бурятия. В Сибир разораването на необработваемите земи е довело до силното намаляване на численноста. Лова на корсака се регулира със специални закони, в които видът се разглежда като обект за търговия с кожи : Русия, Казахстан, Туркменистан, Узбекистан и Монголия. В Русия, Казахстан и Тюркменистан лова се разрешава само от ноември до март, като определени методи на лов са забранени от закона, като например разриването на леговищата, опушването, заливането с вода и използването на отровни примамки. В плен корсака лесно привиква към човека. В 17 ти век в Русия са били популярни като домашни животни.
Афганска лисица, Blanford's fox ,Vulpes cana, Blanford, 1877
![]()
Афганската лисица, наричана още белуджистанска, бухарска, лисица на Бланфорд, се среща в източните региони на Близкия Изток до Афганистан. Описана е за следните страни: Афганистан, Египет / Синай/, Иран, Израел, Йордания, Оман, Пакистан, Суадитска Арабия, Туркменистан, Обединените Арабски Емирства и Иемен. Основната част на ареала е Източен Иран, Афганистан и северозападен Индустан. Обитава полупустинните степи и планините на височина до 2000 м н.в., безлюдни планински местообитания, скалисти склонове, каньони и стръмни скални скатове. Зоолозите считат, че в исторически план видът е избягал от жарките пустинни условия, но сега са срещани такива лисици и близо да Мъртво море, където през летния период има екстремални температури. Виждани са и в селскостопански региони, където се отглеждат грозде, дини, пъпеши и други плодове. В Аравийската пустиня разпределнието на лисиците не е ограничено от достъпа до вода, тъй като те си я набавят от храната. Благодарение на това в Израел лисиците населяват най-сухите и горещи области. Най-високата плътност на популацията е регистрирана в Юдейската пустиня на височина 100 – 350 м под морското равнище.
Филогенетичен анализ, основан на мтДНК, определя афганската лисица и фенека като близки таксони, които формират монофилетичен клон, отличаващ се от останалите същински лисици. Секвенционната разлика между двата вида показва, че те се отделят като видове преди 3-4 млн. години, което се свързва с появата на пустинните региони в Средния Изток и Африка.
![]()
Афганската лисица е малка лисица с височина на холката 30см, дължина на тялото – 40-50 см, дължина на опашката 33-41 см, височина на ухото – 9см., тегло 1,5-3 кг. Зимната окраска на козината е кафяво-сива с по тъмна ивица на гърба, която минава и по дългата и пухкава опашката. Афганската лисица е описвана като вид с котешка външност и поведение. Тази малка лисица има къса, стройна муцуна, дълга, пухкава опашка, която е равна по дължина на тялото на лисицата, с дълги и тъмни основни косми. Ушите са големи, с помощта на които не само имат добър слух, но и разсейват излишната топлина в горещо време. Острата и стройна муцунка има отличителна черна линия от очите до горната устна. За разлика от другите пустинни лисици, възглавничките на лапите не са покрити с косми, които да ги защитават от горещия пясък. Мъжките и женските много си приличат. Зимната козина е мека и пухкава, състояща се от косми с рижаво-кафяв цвят, със сив оттенък и отделни черни косми, което създава един общ кафявосив изглед на окраската с черна ивица, минаваща по гърба до върха на опашката. Корема и гърлото са светли, белеещи. Крайчето на опашката е обикновено тъмен, но макар рядко може да бъде бял. В някой области на ареала цвета на козината може силно да варира от тъмен, почти черен до риж. Големите уши са покрити с редки косми и имат еднороден тъмен кафяв цвят.
![]()
Афганската лисица е всеядна. Основно се храни с малки бозайници и насекоми, така също и плодове . Предимно си набавя храна самостоятелно, рядко по двойки. Нощно животно е, избягва появата през деня. Приспособена е да се катери по скали, затова и лапите са малки и неокосмени. Лесно прескача от един скален склон на друг и се катери по почти отвесни скални склонове. Не са виждани да копаят дупки за леговища, както другите лисици, вместо това използват естествени пукнатини, дупки в скалите и склоновете, за да си направят бърлога.
Моногамни животни са с определена територия. Брачния период е декември – януари. Бременността продължава 50 - 60 дена. Раждат се от 1 до 3 малки, кърменето продължава 30-45 дена. На около 2 месечна възраст започват да се хранят с родителите си и остават с тях до есента. Полова зрялост при тях настъпва на 10 - 12 месечна възраст. Предполага се, че в свободно състояние живеят около 4 - 5 години.
Афганските лисици са ловени заради кожите, които също са били обект на търговия. Основна причина за намаляване на популацията е загубата на хабитати и конкуренцията им с червената лисица. В Израел е забранен лова и отстрела на тези животни, а отглеждането в плен изисква специално разрешение. В Оман и Йордания също са защитени от закона. Видът е включен в червената книга на IUCN като слабо засегнат.
Полярна лисица , Обыкновенный песе́ц, Аrctic fox, Vulpes lagopus , Alopex lagopus Linnaeus, 1768
![]()
Полярната лисица се класифицира в род Vulpes, не рядко в род Canis, най-често обаче е отделяна в отделен род Alopex. Родствено най-близка до нея е американската бързонога прерийна лисица Vulpes velox и лисицата джудже V. Macrotis, макар последните две да не се срещат в тундрата.
Името и vulpes означава лисица, а видовоспецифичното lagopus произлиза от древногръцкото lagos - заек и pous – крака, в съответствие на подобието на лапите им на тези на полярните зайци.
![]()
Полярната лисица е разпространена в района Северния полярен кръг, по крайбрежието и на островите на Северния ледовит океан, в тундровата и лесотундровата зона. Срещат в цяла Арктика, включително Русия, Канада, Аляска, Гренландия и Свалбард, както и в субарктически и алпийски зони, като Исландия и планинските части на Скандинавия. В Русия е типичен представител на фауната на тундрата и лесотундрата. Зимно време, полярната лисица навлиза на юг до Финландия, южната част на Прибалкалието. Виждана е в райони близки до Северния полюс, мигрирала по леда. Южната граница през 20век се мести на север, поради намаляване на ареала на разпространение. По време на последното заледяване, полярната лисица е имала по-широко разпространение по ледниците и нейни фосили се намират в плейстоценски находища в по-голяма част от Европа и Сибир. Филогенетични изследвания, основани на микросателитни локуси и мт ДНК, показват, че популацията на полярната лисица е панмиктична и притежава висока хетерозиготност, както на континента, така и на островите, в Северна Америка, Европа и Азия / Сибир/. Няма съществени генетични различия и характерната ниска хетерозиготност за островните представители. Това се обяснява с няколко причини. Първо по време на заледяванията, видът не се е ограничавал в отделни рефугиуми, а напротив е бил повсеместно разпространен и освен това популацията му е била доста по разширена, във връзка със подходящите условия за живот на вида. При затоплянето, видът е бил ограничаван на север, където освен това е и изтласкван в сегашно време от обикновената лисица, която се конкурира с него за любимата му храна – лемингите. Второ, този вид е способен на големи миграции, за набавяне на храна, като лесно се придвижва по леда, затова реално няма конкретно изолирани отделни, генетично различаващи се субпопулации, дори и на островите. Липсата на генетична структура на популацията на полярната лисица е уникална за групата на канидите.
![]()
Неголяма лисица, с дължина на тялото 50—75 см, дължина на опашката 25—30 см, височина на холката 20—30 см, средно тегло 3,5 кг, максимална – до 9 кг, женските – до 3 кг. За разлика от другите лисици, тялото на полярната е по - ниско, муцуната е по-къса, ушите са закръглени и едва се показват от зимната козина, което предпазва тяхното измръзване. Името и, „зайча лапа” идва от гъстата козина по възглавничките на нейните стъпала, която помага да се ограничи загубата на топлина при ходенето по леда и подобрява сцеплението. Имат сравнително къси крака и голяма пухеста опашка.
Полярната лисица има много дебела и мека зимна козина с гъст, пухест подслой и дълги предпазващи косми.Тази лисица е единствения представител на истинските лисици, който има изразен сезонен диморфизъм на окраската. Бялата форма през зимата има чисто бяла козина, а през лятото мръсно кафява. При синята форма, през зимата има тъмна окраска: от пясъчно и светло кафяво до тъмно сиво със сини отблясъци, дори кафяво със сребърни отблясъци. Сините лисици се срещат при всички популации, но на континента са редки, а на някои острови, напротив, преобладават.
Морфотиповете се определят генетично от един локус, като белите са рецесивни. Сините морфотипове съставляват по-малко от 1% от популацията на континента, 25-30% от популацията на Феноскандинавия и 65-70% от популацията в Исландия. В крайбрежните райони и на островите сините морфотипове могат да достигнат до 100% от местната популация. Между самите морфотипове има вариации в окраската, които не зависят от локуса за окраската.
Описвани са 8, дори 10 подвида, но ние ще се спрем на 4 най-вероятни:
— A. l. lagopus (най-често срещаната).
— A. l. semenovi (остров Медний, Командорските острови, Русия).
— A. l. beringensis (Беринговите острови, Командорските острови, Русия).
— A.l. pribilofensis (острови Прибилов, Аляска).
![]()
Полярната лисица си набавя храна самостоятелно. Когато има храна в излишък, складира за по късно. Полярните лисици се хранят леминги, зайцевидни, птици и техните яйца, мърша и растения. Най-важната сред тези храни са лемингите. Семейство лисици може да изяжда десетки леминги всеки ден. През април и май полярните лисици ловуват и малките на пръстеновия тюлен, които са скрити в снежни леговища и са относително беззащитни. Когато обичайната им плячка е оскъдна, полярните лисици събират остатъците от храната на по-едри хищници, като бялата мечка, въпреки че самата полярна лисица е сред плячката на бялата мечка.
Полярната лисица има редица адаптации към арктическия климат. Козината и има най-добри топлоизолационни свойства от всички животни и индивидите нямат нужда при каквито и да е условия да увеличават нивото на метаболизма си, за да поддържат хомеотермия. Променят цвета на окраската през различните сезони, като така увеличават приспособителния си потенциал и камуфлажната си способност. Окосмяването на лапите също спомага за предпазване от измръзване. Малките уши, късата муцуна и способността да редуцират периферното си кръвообращение, също спомага за намаляване на разсейването на топлината. През есента теглото им може да се увеличи до 50%, като основно се отлагат мазнини за изграждане на топлоизолационен слой и енергиен резерв.
Полярните лисици живеят на брачни двойки и заедно отглеждат потомството. През първите три седмици след раждането на малките, когато те бозаят и майката рядко напуска бърлогата, мъжкия се грижи за набавянето на храната за двамата. Когато малките започнат да се хранят с месо, тогава и двамата родители участват в подсигуряването на храна поравно. Помощ може да оказват останали в семейството от предишното котило, вече пораснали малки. Женските помощници напускат семейството, преди малките да са навършили 8-10 седмична възраст. На остров Медний са наблюдавани постоянни семейни групи с повече от 6 възрастни. На други острови също са наблюдавани сложни социални системи.
Полярните лисици са строго териториални по време на брачния период, като обичайно едно семейство да обитава една бърлога, но понякога бърлогата може да се подели и с други близкородственици.
Територията на едно семейство варира от разпространението на лемингите, любимата храна на тези лисици, от 15 до 36km²; на островите 5 – 21km², в зависимост от територията на самия остров, на Свалбард 10 – 125km². Обичайно е маркирането на територията с урина. Вокализацията и позирането са част от сложните сигнализационни системи на общуване.
В Аляска са наблюдавани сезонни миграции, когато индивидите напускат зимните си бърлоги и мигрират към крайбрежието, като се връщат обратно късна зима, ранна пролет. Големи миграции са наблюдавани в Канада, Феноскандинавия и Русия. Те могат да бъдат причинени от липсата на храна, като сериозен спад на популацията на лемингите. Най-дългата регистрирана миграция е на мъжки индивид намерен на 2300 км от първоначалната точка.
Брачния период е от февруари до май. Малките се раждат от април до юли, след 51-54 дневна бременност. Малките тежат при раждането си 80-85 г и са безпомощни. Броя на малките варира на различните места от 2 до 6, макар на някой места да са регистрирани и повече от 19 малки.Големината на малките принципно варира и зависи от периода на наличие на храна, при по-висока численост на гризачи в района се раждат по-големи малки и обратно. Способността на полярните лисици да произвеждат голямо потомство е обусловено от достъпа до голяма и сигурна бърлога. Първоначалната функция на брачното леговище е да подсигурява защита срещу други хищници. Бърлогата представлява сложна подземна система от проходи и дупки, като най-широката част е мястото за раждане. Този сложен комплекс може да има повече от 150 входа.Добрата бърлога е разположена над вечно замръзналата почва, на места където се натрупва по-малко сняг и обикновено е с южен изглед. В Канадската тундра са описани бърлоги на приблизително 330 години. Някой леговища се използват регулярно, всяка година, докато други от време на време. Отглеждането на малките е ограничено в периода без сняг – юни до септември. Лактацията продължава 8-10 седмици. Достигат полова зрялост на 10 месечна възраст. Продължителност на живота в природата до 3 години, макар да е регистрирано и до 11 годишна възраст.
![]()
Изобилието на полярната лисица се изменя циклично, едновременно с размера на популацията на лемингите. Тъй като лисиците се размножават много бързо и често умират млади, броят на популацията не се влияе силно от лова. Въпреки това те са изчезнали в много области, заселени от хора.
Полярната лисица губи територии от обикновената лисица. В миналото вълкът е ограничавал числеността на обикновените лисици, но с почти пълното изтребване на вълците, броят им е нарастнал. Полярната лисица сама става жертва на по-големи от нея хищници, освен вълка, бялата мечка, росомохата, орли, бели сови.
Полярната лисица е обект на добива и търговията с кожи, заради ценната си кожа. Особено високо се цени кожата на синята лисица, която се и отглежда в плен. На островите лисиците се отглеждат свободно. Ферми за отглеждане на полярни лисици има в северните и средни ширини на Америка, Европа и Азия.
Лова е бил дълго време главен летален фактор за популацията на полярната лисица. В Северна Америка за периода от 1919 – 1984 са избивани приблизително 40 000 – 85 000 лисици годишно. В Канада 10 000 – 68 000 лисици годишно. В Аляска за периода от 1925 до 1962 от 3900 до 17 000 лисици годишно, като по късно добива спада на 1000 – 2000 годишно.
В Сибир лова за кожи достига в някой години до 100 000 лисици годишно, като през някой години на 70 те, 35 – 59% от популацията е била убивана всяка година. Намаляването на популацията в Сибир през последните няколко деситилетия е очевидно, макар спада в цената на тази кожа и пробивите в съветската търговска система има ефекта върху отслабване на натиска върху популацията на лисицата от човека.
Важността на кожата на полярната лисица има дълга история. Йорданес в Гетика описва римляните, че са облечени в тъмно сини кожи, закупени от свеите / германско племе, населявало днешна Швеция/, вероятно закупени преди това от Screrefennae / саамите/. Икономиката на инуитите е тясно свързана с обилието от полярни лисици. Кожите на лисиците са били законно платежно средство, наред с агнешките и с някой други продукти в Исландия през Средните векове.
![]()
Полярната лисица остава единствения, най-важен обект на лов в Арктика. Местните жители винаги са използвали нейната изключителна кожа и с основаването на кожарската промишленост, полярната лисица бързо става важен източник на доходи. Днес капаните и отстрела са основни ловни похвати. Поради високата си репродуктивна способност, полярните лисици успяват да задържат сравнителна стабилност на популацията, въпреки натиска от страна на човека. В някой региони са избивани повече от 50% от популацията на вида, въпреки че това не е позволено от закона, както е във Феноскандинавия. Лова на полярни лисици напоследък намалява, поради спада на цената на кожата и алтернативни източници на доходи. В Юкон, например, общия доход от кожарската индустрия спада от 1,3 млн долара през 1988г на по-малко от 300 000 долара през 1994г.
![]()
В по-голямата част от ареала си, полярната лисица не е защитена. Вида и неговите леговища са защитени законово в Швеция от 1928г, Норвегия от 1930г, Финландия от 1940г. В Европа полярната лисица е приоритетен вид в действията на Actions by the Community relating to the Environment (ACE). В Норвегия / Свалбард/, Гренландия, Канада, Русия и Аляска, лова на тези лисици е ограничен и се разрешава само в определено време. Видът е включен в червената книга на IUCN като слабо засегнат.
Лисица, Червена лисица, Ры́жая лиси́ца, Red fox, Cross fox, Silver fox, Vulpes vulpes, Linnaeus, 1758
![]()
Лисицата, наричана още червена лисица или обикновенна лисица, е най-често срещаният вид лисици и има най-широк ареал от всички сухоземни хищници. Тя е най-големият вид в рода Лисици (Vulpes) и обитават цялото Северно полукълбо от Северния полярен кръг до Северна Америка, Централна Америка и Азия. Нейния ареал се разширява заедно с този на човека. Интродуцирана е в Австралия през 18 век, където се оценява като вредна за местните бозайници и птичите популации на континента. По тази причина е включена в списъка на „100 те световни най-отрицателни инвазивни видове”. Не се среща в Исландия, арктическите острови и някой части от Сибир, както и в същинските пустини. Европейските подвидове са интродуцирани в източните САЩ и Канада през 17 век, където се смесват с местните подвидове. Преди се е считало, че северноамериканската лисица Vulpes fulva е отделен вид, но сега се приема, че е палеарктичната червена лисица.
![]()
Червената лисица произхожда от малък предшественик от Евразия по време на среден вилафранкски период и колонизира Северна Америка кратко след уисконсинското заледяване. Предполага се, че нейния предшественик е или Vulpes alopecoides или V. chikushanensis / Китай/. Най-ранни фосили от V. vulpes са намерени в Барани, Унгария и датират отпреди 3,4 и 1,8 млн.год. Предшествениците са били по- малки от съвременните видове. Сред същинските лисици, червената лисица демонстрира по-прогресивно развитие в насока към хищничество. Освен по размера си, червената лисица се отличава от другите лисици по способността си бързо да се адаптира към промените на околната среда.
Червените лисици колонизират Северна Америка на две вълни: по време или преди Илинойското заледяване и по време на Уисконсинското заледяване. В далечния север фосили на червени лисица са открити в Сангамонски отлагания в област Феърбанкс и Медисин Хат. Фосили от времето на Уисконсинското заледяване са намерени в 25 находища в Арканзас, Калифорния, Колорадо, Айдахо, Мисури, Ню Мексико, Тенеси, Тексас, Вирджиния и Уайоминг. Генетични изследвания , основани на сравнителен анализ на фрагменти мтДНК, разделят лисиците на два основни хаплотипа и респективно два основни клона – Холарктичен клон, включващ Европа, Азия, Аляска и западна Канада; и Неарктичен клон, включващ останалата част на Северна Америка. Тези изследвания показват, че по време на Уисконсинското заледяване е имало два основни рефугиума в Северна Америка. Северния рефугиум се е намирал в Аляска и западна Канада, като в него са влизали подвидовете V. v. alascensis, V. v. abietorum, V. v. regalisand V. v. Rubricosa. Южния рефугиум се е намирал в предпланинските паркове и алпийските ливади на Скалистите планини, Каскадите и Сиера Невада.Този клон по късно се разделя на всички останали популации след последното заледяване и има специфични екологични и физиологични адаптации.
![]()
Фиг.1 Пълна медиан – свързваща мрежа, основана на фрагмент от мтДНК с дължина 354 bp на 220 лисици.
![]()
Диаграма на разселването на червената лисица от Стария свят в Новия по време на плейстоценските заледявания. Подобно е разселването на лисиците в Япония на островите Хокайдо и Хоншу, на които лисиците се отличават помежду си хаплотипно.
В Европа филогенетични изследвания, основани на фрагменти мтДНК показват висока хетерозиготност и ниска генетична структура на популацията на лисицата.
Описани са 44 подвида на червената лисица, макар някой да са спорни. Най-общо се делят на големи северни лисици и малки южни лисици от Азия и Средния Изток:
— V. v. abietorum ( Стюарт Лейк, Британска Колумбия, Канада)
— V. v. aegyptiaca (Eгипет)
— V. v. alascensis ( Андреафски, Аляска, САЩ)
—V. v. alpherakyi (Геок Тепе, Араиск, Казахстан)
— V. v. anatolica (Смирна, западна Мала Азия, Турция)
— V. v. arabica (Мускат, Оман)
— V. v. atlantica (Атласките планини, Мидия, Алжир)
— V. v. bangsi (L’Anse au Loup, Бел Айл , Strait of Belle Isle, Лабрадор, Канада)
— V. v. barbara (Варварски бряг, северозападна Африка)
— V. v. beringiana (бреговете на Беринговия проток, североизточен Сибир)
— V. v. cascadensis ( планинската верига Каскади, окръг Скамания, Вашингтон, САЩ)
— V. v. caucasica (близо до Владикавказ, Кавказ, Русия)
— V. v. crucigera ( Тюрингия, Германия)
— V. v. daurica (Карангой, 45км западно от Троицкосавск, Сибир)
— V. v. deletrix ( Залива Сейнт Джордж, Нюфаунленд, Канада)
— V. v. dolichocrania ( Сидеми, южно Усури, югоизточен Сибир)
— V. v. flavescens ( северен Иран)
— V. v. fulva ( Вирджиния, САЩ)
— V. v. griffithii ( Кандахар, Афганистан)
— V. v. harrimani ( остров Кодиак, Аляска, САЩ)
— V. v. hole ( близо до Амой, Фукиен, Южен Китай)
— V. v. ichnusae (Сарабус, Сардиния, Италия)
— V. v. induta ( Пила, Кипър)
— V. v. jakutensis ( тайга, южно от Якутск, Източен Сибир)
— V. v. japonica ( Япония)
— V. v. karagan ( киргизките степи, Киргизия, Русия)
— V. v. kenaiensis (Полуостров Кенай, Аляска, САЩ)
— V. v. kurdistanica ( долината Гелск, североизточна Турция)
— V. v. macroura (планините Уосатч, близо до Голямото солено езеро, Юта, САЩ)
— V. v. montana ( Хималаите)
— V. v. necator ( Уитни Медоу, близо до планината Уитни, окръг Тюлери, Калифорния, САЩ)
— V. v. ochroxantha (Аксай, Семиречие, Източна Русия, Тюркестан, Киргизия)
— V. v. palaestina ( Рамлех, близо до Ramleh, близо до Яфа, Окупирана Палестинска територия)
— V. v. peculiosa ( Корея)
— V. v. pusilla ( Соления хребет, Пинджаб, Пакистан)
— V. v. regalis ( Елк Ривър, окръг Шърбърн, Минесота, САЩ)
— V. v. rubricosa (Дигби, Нова Скотия, Канада)
— V. v. schrencki (Сахалин, Русия)
— V. v. silacea (близо до Силос, Бургос, Испания)
— V. v. splendidissima (северни и централни Курилски острови, Русия)
— V. v. strepensis (степите, близо до Херсон, Русия)
— V. v. tobolica (Обдорск, Тоболск, Сибир)
— V. v. tschiliensis (Пейпинг, Чихли, Североизточен Китай)
— V. v. vulpes (Швеция)
Въпреки името си, червената лисица има морфотипове и в други цветове, включително албиноси и меланистични форми. Албиносите се отличават от полярните лисици, главно по размера си, големината на ушите и по-издължената муцуна.
Цветови вариант | Изглед | Описание |
Aмбър | ![Vulpes vulpes (Amber fox) fur skin.jpg]()
| |
Чернокафяв
/ въглищарка/ | | Меланистична форма на евразийската червена лисица. Има черно кафява или черна козина със светло кафяв отенък. Кожата обикновено е смесица от различен отенък на сребърното. Червеникавата козина или липсва напълно или присъства с редки кичури. |
Кръст / крос/ | ![Vulpes vulpes (cross fox) Norway & Canada.jpg]()
| Козината има по-тъмна козина от нормалната.Задницата и долната част на гърба са тъмнокафяви до тъмно сиви с различна степен на сребристото на защитни косми. Кръстът на раменете е черен или кафяв, по рядко бледо сребристо. Краката и главата са кафяви. |
Сива | | Задницата и гръбнака са кафяви или сиви с бледо жълтеникави ивици на защитните косми. Кръстът на раменете е кафяв, ръждиво кафяв или червеников кафяв. Краката са кафяви. |
Платинена | ![Vulpes vulpes (Platinum fox) fur skin.jpg]()
| Отличава се от сребистия морфотип по нейната бледо сребриста, почти сребристо бяла козина със синкави отенъци. |
Червена | ![Red fox fur skin (Sweden).jpg]()
| Типичната окраска за вида |
Самсон | ![Vulpes vulpes Mutation.jpg]()
| Отличава се със своята вълнена кожа, на която липсват защитни косми. |
Сребърна | ![Vulpes vulpes - silver fox fur skin.jpg]()
| Меланистична форма на Северно американската червена лисица, която се внася в Стария Свят , заради търговията с кожи. Характеризира се с чисто черна окраска, с варираща смесица на сребристото / покриващо 25 – 100% от площта на кожата/ |
Червената лисица е най-голямата от всички лисици. Дължина на тялото 60—90 см, дължина на опашката 40—60 см, тегло 6—10 кг. Окраската и размерите на тези лисици са различни в различните местности, като са описвани 40 – 50 подвида, без да се броят най-малките форми. На север лисиците стават по-големи и светли по окраска, а на юг по- малки и по-тъмни. В северните райони и в планините често се срещат черно-кафяви и други меланистични форми на окраска. Най-разпространена е червено – рижата окраска: ярко риж гръб, бели гърди и корем, тъмни лапи. Често имат тъмни ивици на лопатките и гърба, така че напомнят кръст. Основни отличителни черти са тъмните уши и белия край на опашката. Външно лисицата представлява животно със среден размер, изящно сложено тяло на невисоки, тънки крайници, с издължена муцунка, остри уши и дълга пухеста опашка. Линеенето започва в края на февруари - март и завършва в средата на лятото. Веднага след това започва израстването на зимната козина, която напълно се формира и носи през ноември и декември. Лятната козина е по-рядка и къса, докато зимната е по-дълга, гъста и пухеста. Лисиците се отличават с големи уши – локатори, с помощта на които улавят звуковите колебания, за да уловят и най-малкия звук на плячката. Нейния слух е толкова добре развит, че може да усети мишка под 60 см снежна покривка.
В България има два морфотипа – червена лисица и въглищарка. Червената лисица има рижаво-оранжев или жълто-оранжев гръб и горна част на тялото. Долната част на гърдите и гушката са почти бяло-сивкави. Краката и задната част на ушите са почти черни. Краят на пухкавата рижава опашка е бял. Това е и преобладаващата окраска в Европа. Характерно е, че преходът от сиво към червено окосмяване не е рязък, а става с плавно преливане. Лисиците въглищарки или кюмюрджийки, са срещат често у нас. Техния основен цвят е тъмнокафяв, черен, гърлото е тъмносиво, края на опашката е или бял или черен.
![]()
Червената лисица има бинокулярно зрение, но реагира основно на движение. Слуха е изключително добре развит, като е възможно да чува тетрев, който прелита от едно място на друго от 600 крачки / приблизително 457 м/ разстояние, полета на врани от 250 до 500 метра и писъка на мишка от 100 метра. Движенията на това животно са много гъвкави. Тя е издръжлива и може да се носи в бърз тръс, когато преследва заек или да мине в светкавичен галоп. Плува отлично и може да прави удивителни, координирани и във въздуха, скокове. Това е много подготвен хищник, крайно издръжлив и упорит, при това изобретателен , дързък и умен.
Имат чифт анални торбички, заобиколени от мастни жлези, като и двете се отвеждат с един отвор навън. Във всяка анална торбичка има ферментационна камера с аребни и анаеробни бактерии, които превръщат себума в мирисно вещество, вкючително алифатни киселини. Овалната опашна жлеза е 25 мм дълга, 13 мм широка и е описана да мирише на виолетки. Когато почиват, завират муцунката си близо до тази жлеза.
Голямото разнообразие на окраската и големината на червената лисица е свързано със ширината на нейния ареал и голямото разнообразие на условията, при които живее в отдлените му части. Достатъчно е да се каже, че лисиците заселяват, макар и с различна плътност, всички ландшафтни – географски зони, като започнем от тундрата и субарктическите гори до степите и пустините, включително планинските масиви във всички климатични зони. При това лисицата обитава не само дивата природа, но и културни ландшафти, както и в покрайнините на големи градове като Киев и Варшава. В Лондон, лисицата е обичайна за покрайнините на града, а понякога се появява дори и в централната част на града. Освен това, във времето на активна урбанизация на територии, лисицата намира благоприятна за себе си среда. Нерядко живее в депа за отпадъци, паркове и мазета. Във всички части на своя ареал, лисицата предпочита открити местности, а също така райони, където има отделни насаждения, хълмове, гори, дерета, особено ако зимата няма много дебела снежна покривка. Така от всички климатични зони, най-много лисици обитават степните и лесостепните, а не в планинските.
Принципно лисицата не е мигриращ вид. Случай на миграции са отбелязвани само в тундрата, пустините и планините. Например, една от белязаните в Малоземелската тундра / Архангелска област, Русия/ лисица е била намерена убита на 600 км югозападно. Млади животни, които се разселват, обичайно се намират на разстояние от 2 – 5 до 15 – 30 км от леговището на родителите си . Числеността на лисиците доста се колебае през годините. На нейното състояние влияят такива фактори като количеството гризачи, метеорологичните условия, наличието в популацията на инфекциозни заболявания. В гладните години не само спада плодовитостта на женските и оцеляват по-малко лисичета, но и възникват условия, благоприятни за разпространението на болести, които понякога могат да се разпространят широко. Характерни за лисицата болести са бяс, чума по хищниците и краста.
Лисицата, въпреки че принадлежи към хищниците се храни с доста разнообразна храна. Изследвания върху менюто на лисиците показват, че те могат да се хранят с над 400 вида животни и десетки видове растения. Основната част на менюто са гризачите, главно полевки. Може да се каже, че тяхното обилие и достъпността им в значителна степен влияят на популацията на лисицата. Това е особено важно през зимата, когато лисицата живее основно от лова на полевки, като ги чува под снега и с бързи скокове започва да се рови в снега, като го разхвърля са лапи, за да ги хване. Този начин се нарича мишкуване.
![]()
По големи бозайници, като зайци например, имат по-малка роля в храненето, макар някой лисици да обичат да ги ловят, особено зайчетата, а по време на заешко измиране може да се хранят и с труповете им. По-големи лисици могат да нападнат и сърнета. Птиците в менюто на са така важни, както гризачите, макар че лисицата няма да пропусне случай да хване птица, която се е оказала на земята / от най-малки до по-големи птици като гъски и глухари/, похапват и птичи яйца или нелетящи птиченца. Лисицата похищава и домашни птици, но по наблюдения това тя прави много по-рядко отколкото се твърди. В пустините и полупустинните лисиците често се хранят с влечуги. В Канада и североизточна Евразия, лисици които живеят край реки, сезонно се хранят почти на 100% със сьомги, които са умрели след хвърлянето на хайвера. Лятото лисиците ядат много бръмбари и други насекоми. В гладни времена не отказват и мърша. Растителната храна включва основно плодове, по-рядко вегетативни растителни части.
Индивидуалния участък, който заема една двойка или семейство лисици е длъжен да ги обезпечава не само с достатъчно количество храна, но и с пригодни за устройване на леговища места. Лисиците ровят леговищата си сами, макар понякога да заемат чужди на язовци, полярни лисици, мармоти и други ровещи животни, като ги приспособяват за свои нужди. Най-често лисиците се заселват на склонове и хълмове, където има пясъчен грунт, защитени от дъждовни и други води. Обикновено леговището има няколко входове, които водят в различно дълги тунели към гнедовата камера. Понякога лисиците използват естествени заслони, като пещери, процепи в скалите, хралупи в стари дървета. По принципи леговището се използва най-активно по време на отглеждане на малките, в останалото време лисиците могат да си почиват на поляната или на снега. Ако леговището се зарази с паразити, лисиците правят ново. В полето използват всяко укритие, за да си направят дупка. Синори, каменни огради и дори стари напоителни канали. Има случаи да импровизират някакво леговище само за малките, а възрастните да живеят навън до есента. Лисиците са топлолюбиви животни, а в дълбоките дупки температурата е постоянно 12 – 14 С и така оцеляват през зимата. През слънчевите дни излизат и се припичат до някой дънер на дърво. Леговището им служи по-скоро за убежище, където бързо се скриват при преследване, при дъжд и сняг.
Лисицата е моногамно животно и се размножава веднъж годишно. Брачния период и неговата ефективност зависи от времето и натрупаните мазнини. Има години, когато до 60% от женските остават без потомство. Още през зимата лисиците изберат място за леговище, където ще отглеждат малките и ревностно го пазят. За една женска се случва да се борят и двама трима мъжки. Размножителният период на лисицата варира в зависимост от географския район, който тя обитава. Тези, които са на юг, се размножават през декември-януари; северните популации се размножават през февруари-април; а индивидите, които се намират между южните и северни лисици, се размножават през януари-февруари Лисиците са добри родители. Мъжките имат активно участие във възпитанието на малките лисичета, а освен това се грижат за майката до появата на потомството. Благоустрояват леговището и дори ловят на женските бълхите. Бременността продължава 49 – 58 дни, след което се раждат от 4 – 6 до 12 – 13 малки. Малките се раждат слепи и тежат до 150 грама. Проглеждат и започват да чуват до около 2 седмици и правят първи крачки извън бърлогата до 5 седмици. За малките се грижат и двамата родители, като са изключително внимателни по отношение на опазването на малките и при опасност ги местят в резервно леговище. Подрастващите лисичета от рано разпознават дома си и могат да се намерят на разстояние от него. Кърменето продължава около месец и половина, след което малките се приучават на нормална храна, както и ги обучават в намирането и. След 6 месеца подрасналите лисичета започват да се разселват. Към есента вече са напълно зрели и могат да живеят самостоятелно. Мъжките могат да се отдалечат на 20 – 40 км, женските на 10 -15, рядко на 30 км от дома, в търсене на своя територия и място за свое леговище. Достигат полова зрялост на 8 – 10 месеца. В природата лисиците рядко живеят повече от 7 години, често продължителността на живота не надвишава 3 години. В плен доживяват 20 – 25 години.
Когато се придвижва спокойно, лисицата върви по права линия, оставяйки след себе си ясна пътека от следи. Наплашеното животно, може да бяга много бързо, в галоп и буквално носещо се над земята с опъната опашка.
Лисиците комуникират помежду си с помощта на доста звуци, мимики, пози и с миризми. Около 28 различни категории вокализации са описани.Когато е разгонена или просто в състояние на възбуда, лисицата издава отривист и гръмък лай, а когато се бият, пронизително пищят. Гласа на мъжкия и женската се различават, женската излайва три пъти и завършва с кратък вой, докато мъжкия лае като куче, без да вие.
Много лисици, особено младите, се излежават през деня в полето, особено ако е разположено близо до гората и е богато на гризачи. Преди да легне под някой храст или до пън, лисицата замръзва на място и наблюдава дълго околността за опасност. След това ляга, завива се на кръг и прикрива носа и лапите с опашката. Обичат да почиват в гъсти, обрасли с храсти места, труднодостъпни. Ловуват по различно време, като предпочитат сумрака на ранното утро или привечер, а където са спокойни, могат да излязат на лов и през деня. Когато има възможност от опасност, лисицата е много внимателна и с удивителна способност да се крие от преследване. Именно затова във фолклора на много народи, лисицата се явява въплащение на хитростта и ловкостта. Лисици, които живеят до туристически места, места за отдих и там където е забранен лова, бързо привикват към присъствието на човека, лесно идват да бъдат нахранени и може даже да се научат да просят храна от хората. Предполага се, че лисиците имат развито чувство за магнитно поле.
Лисиците се развиват отлично в регионите, в които няма вълци. Вълците свирепо унищожават лисиците, чакалите също ги подтискат, защото ловуват организирано и на същите ловни места. Кучетата остават най-сериозните врагове на лисиците. Те ги преследват навред. Когато е гладна, лисицата става дръзка и предприема рисковани набези. Често, когато храни малките си, става изключително нахална и непредпазлива. Тогава нарастват и нападенията над домашните птици в селата. При това те могат да са на километри от леговището. Лисицата може да носи голяма кокошка с часове, за да нахрани лисичетата си.
Лисицата има голямо стопанско значение като ценен ловен обект, а също така регулатор на числеността на гризачите и насекомите. При това загубите от домашни животни са в пъти по-малки от ползите, които лисиците носят. Ловувани са за спорт, като във Великобритания например ловът на лисици е традиция, в която се включват кучета и преследвачи на коне. За добив на кожи, лисици се отглеждат в плен. В края на XIX век, изкуствено е въведена породата на сребристо – черните лисици. Благодарение на селекция при тази порода е значително подобрено качеството на козината, а защо така се отглеждат различни кожени породи на нейна основа: платинена, бакурианска, дакотска и др.
През 1959 г от директора на Института по цитология и генетика към РАН, Новосибирск, е започнат дългогодишен експеримент за одомашняване на сребристо – черни лисици. В процеса на експеримента се отбират за размножаване само доброжелателно настроени към човека животни. В резултат се създава популация от одомашнени сребристо – черни лисици, при които има различия във физиологията, морфологията и поведение спрямо дивите им родственици. Домашните лисици се държат подобно на домашните кучета: при появата на човека махат с опашки, вият, изискват внимание, галят се. Наскоро учените продадоха на САЩ, Германия и Нидерландия такива одомашнени лисици. А през интернет на специален сайт, всеки може да си закупи такова животно за приблизително 8 000 $. / снимки и информация на http://bigpicture.ru/?p=352553/.
![]()
Рижата лисица е много разпространен персонаж във фолклора на различни страни от света. В европейските страни тя, като правило, въплащава хитростта и коварството, встъпвайки в разнообразни роли : от вероломен негодяй / както е в редица руски приказки/ до умен съветник. Един от най-известните лисичи персонажи е Лисицата Ренар – персонаж на класическа средновековна поема „ Роман за лисицата”. В месопотамската митология, лисицата е свещенно животно. Тя служи на богинята Ки като ловец. Във Финландия лисицата е символ на хитрост, но не и зло. В японската митология съществуват лисици - духове кицун / подобни на върколаците, но лисици/, които се преобразяват в човешки облик. Те притежават огромни знания и владеят магии. По късно кицун стават известни в литературата, киното и видео игрите. Духове, подобни на кицун, фигурират също така в китайските и в корейските митове. В гръцката митология, Тевместката лисица е била гигантска лисица, дете на Ехидна, която е нападнала Беотия. В келтската митология лисицата също е била символно животно. В Котсуолд, вещиците се преобразявали в лисици, за да крадат масло от съседите.Лисицата присъства в много литературни произведения, анимации, кино филми, песни, аниме, компютърни и видео игри, уеб комикси. В хералдиката, лисицата е символ на проницателност, хитрост и прозорливост. Известно е ловуване на лисици от 4 век пр. Н.е. Лова на лисици с кучета е бил част от европейската култура до 11 век и е било широко разпространен от британските колонизатори.
Днес влияние на популацията на червената лисица оказват загубата на хабитати и преследването за лов и кожи. Търговията с кожи на тези лисици е най-голяма през 20 век. Общия световен оборот на кожи на диви червени лисици за 1985 – 1986 е бил 1,543,995 кожи. Кожите на 45% от уловените диви американски лисици се изчисляват на стойност $50 млн.
Видът е включен в червената книга на IUCN като слабо засегнат.






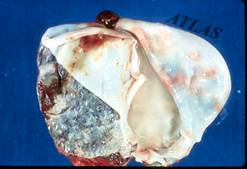
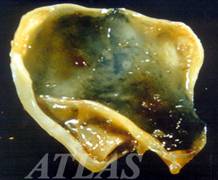


















































































 Дължина на тялото: 71см; Размах на крилете: 157см.
Дължина на тялото: 71см; Размах на крилете: 157см.



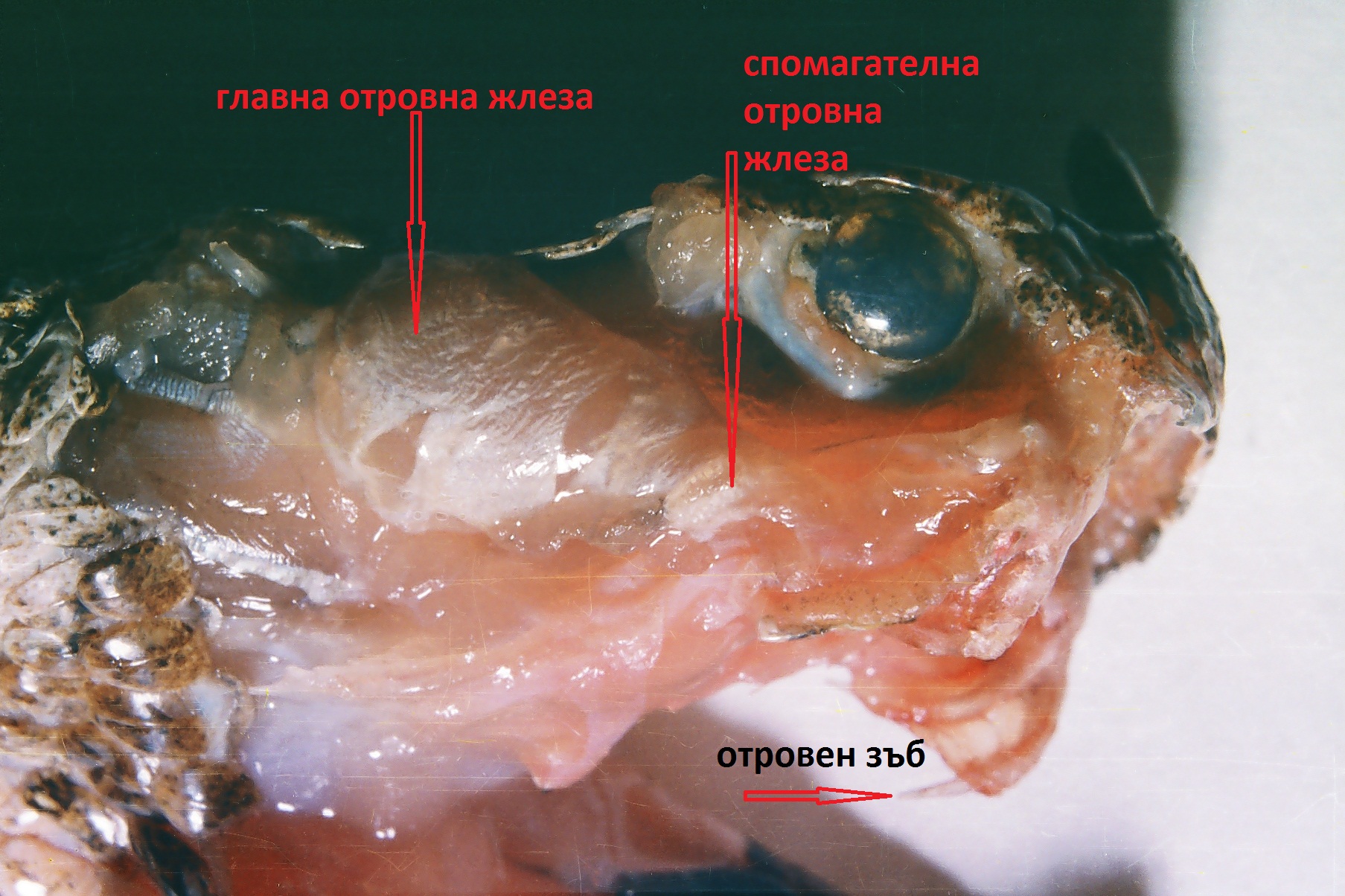
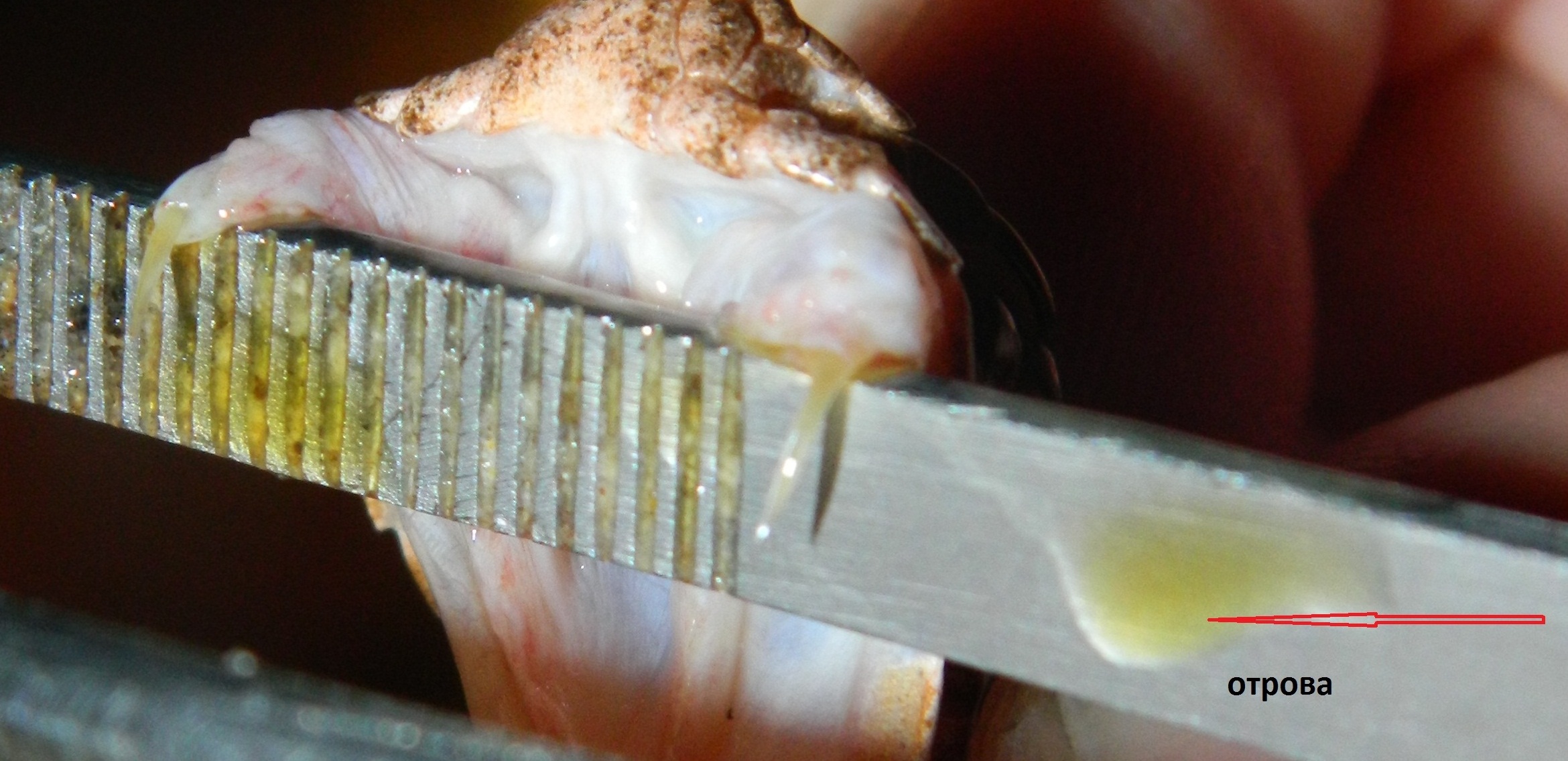


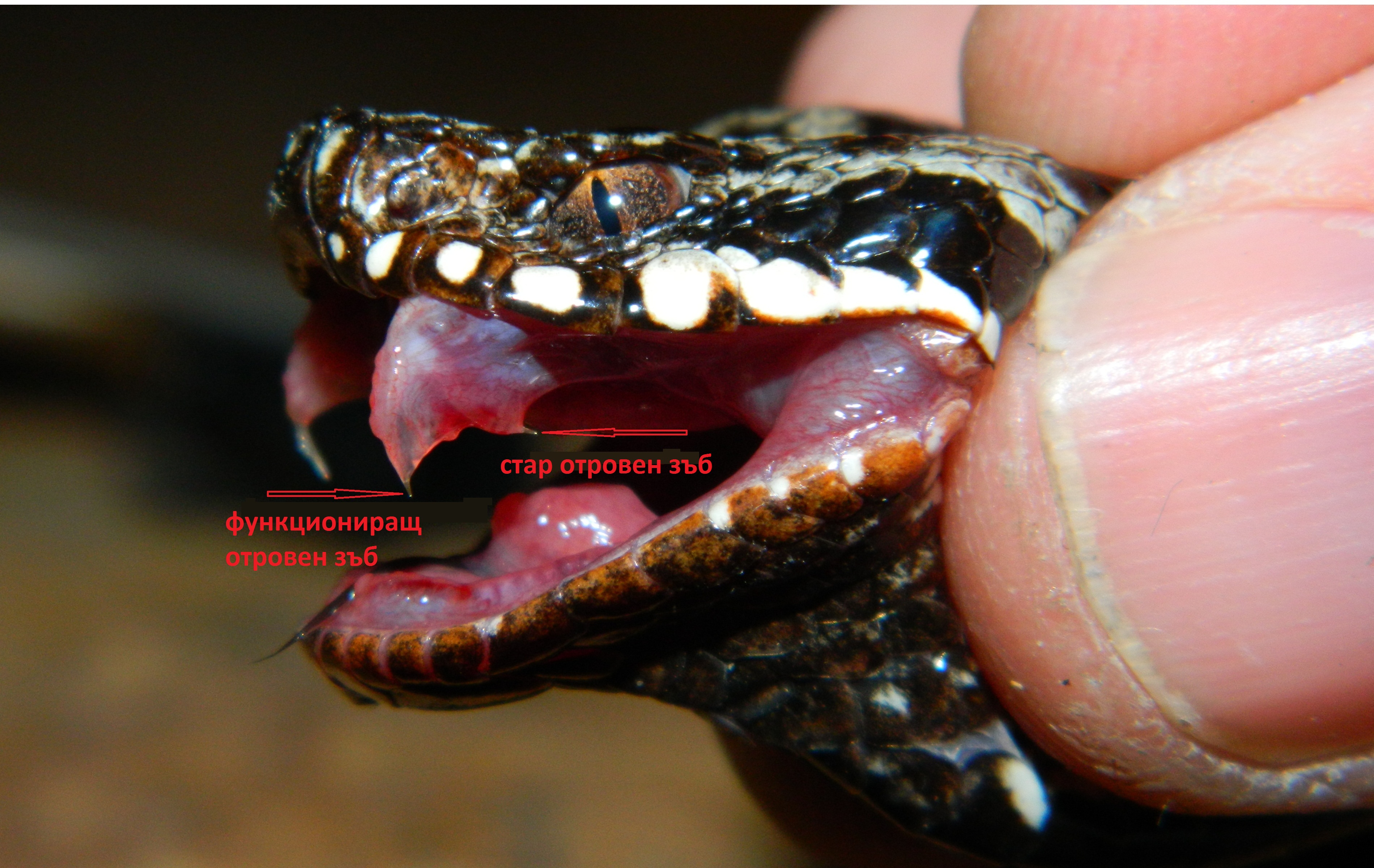
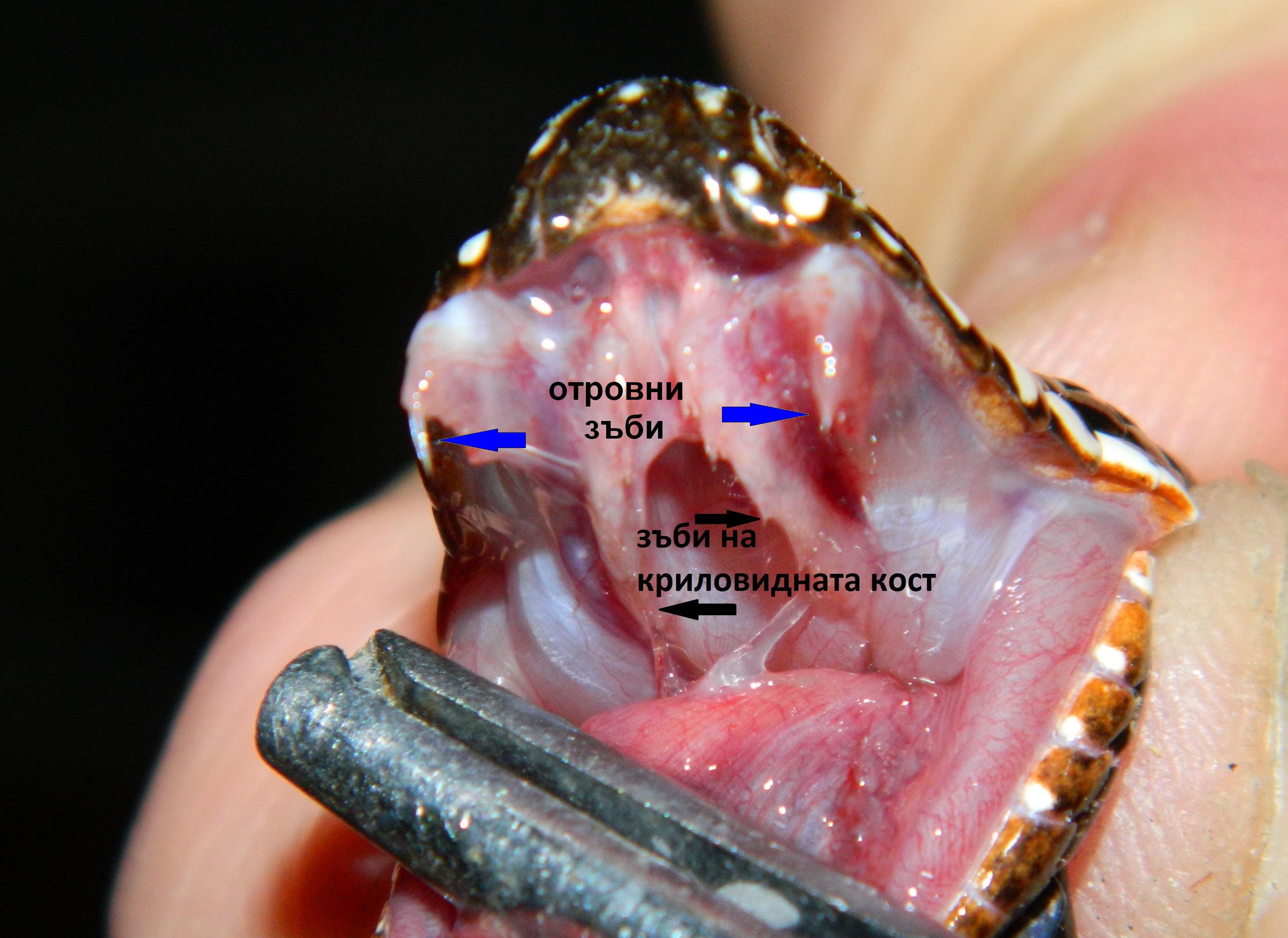




















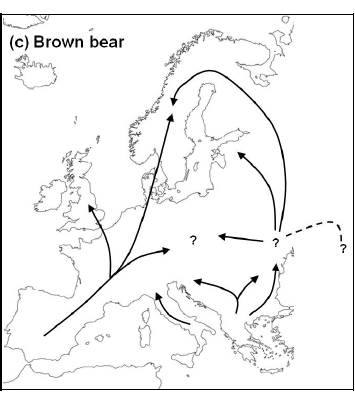





















 Тази неголяма мечка е уникална сред своите родственици, поради основното им меню насекоми – мравки, термити и др. Бърнестата мечка обитава горските и тревисти райони на Индийския полуостров. Има дълга, пухеста козина, с удължени около рамената участъци, оцветена в черно. Рядко се срещат в канелен, червеникав цвят на козината. Отпред на гърдите имат бяло или кремаво петно в U’ или Y’ форма. Характерна е източната муцуна в по-блед цвят. Формата на муцуната, голите бърни и липсата на горни резци са адаптация за хранене с насекоми. Предните лапи са извити навътре и имат дълги, леко закривени нокти, предназначени за ровене и копаене.
Тази неголяма мечка е уникална сред своите родственици, поради основното им меню насекоми – мравки, термити и др. Бърнестата мечка обитава горските и тревисти райони на Индийския полуостров. Има дълга, пухеста козина, с удължени около рамената участъци, оцветена в черно. Рядко се срещат в канелен, червеникав цвят на козината. Отпред на гърдите имат бяло или кремаво петно в U’ или Y’ форма. Характерна е източната муцуна в по-блед цвят. Формата на муцуната, голите бърни и липсата на горни резци са адаптация за хранене с насекоми. Предните лапи са извити навътре и имат дълги, леко закривени нокти, предназначени за ровене и копаене.























 Наричана на староанглийски още бруун или бруин, което също означава кафява, в Дивия Запад, мечките гризли ги наричали Олд Ефрейм /"Old Ephraim" / или Мокасиновия Джо /"Moccasin Joe"/, латинското наименование идва от латинската дума мечка - ursus и старогръцката дума мечка - arctos. Кафявите мечки са широко разпространени в голяма част от Северна Америка и Евразия в гористи и планински региони. Този вид е национално животно на много щати в Северна Америка и страни в Европа и Азия.
Наричана на староанглийски още бруун или бруин, което също означава кафява, в Дивия Запад, мечките гризли ги наричали Олд Ефрейм /"Old Ephraim" / или Мокасиновия Джо /"Moccasin Joe"/, латинското наименование идва от латинската дума мечка - ursus и старогръцката дума мечка - arctos. Кафявите мечки са широко разпространени в голяма част от Северна Америка и Евразия в гористи и планински региони. Този вид е национално животно на много щати в Северна Америка и страни в Европа и Азия.
























 “..Сам Дядо Коледа - дребничък стар симпатяга,
“..Сам Дядо Коледа - дребничък стар симпатяга, 













































































































































 При определени условия, които все още се изясняват, жълтите змиорки напускат континенталните си местообитания и започват своята дълга хиляди километри размножителна миграция. По време на тази миграция настъпват редица приспособителни физиологични и морфологични изменения. Тялото променя оцветяването си и в крайната фаза на промяна, когато вече навлизат в океана се превръщат в сребърни змиорки. Очите им се уголемяват и придобиват тъмно син цвят, променят се и пигментите на ретината им. Жълтата змиорка използва преимуществено мирисните си рецептори, докато сребърната – зрението и то специализирано за улавяне на светлина и на значителна дълбочина. Перките нарастват, кожата става дебела, страничната им линия се обособява доста добре и става видима. Настъпват и редица физиологични промени, които са свързани с промяната на химичния състав и солеността на водата, така че от придънен вид, криещ се в цепнатини и заравящ се в тинята, тя става активно плуващ, пелагичен вид. Стената на плавателния и мехур се увеличава и ефективността на газовата обмяна се увеличава многократно. Натрупват се около 20% резервни мазнини, запаси за дългото пътуване и половото узряване. Всички тези промени имат една цел – сребърната змиорка е приспособена за дълго, продължаващо хиляди километри плуване, за да може да достигне до местата за хвърляне на хайвер. По време на тази миграция змиорката спира да се храни и узрява полово, така че когато пристигне на мястото на размножаване, тя е готова да хвърли хайвера си.
При определени условия, които все още се изясняват, жълтите змиорки напускат континенталните си местообитания и започват своята дълга хиляди километри размножителна миграция. По време на тази миграция настъпват редица приспособителни физиологични и морфологични изменения. Тялото променя оцветяването си и в крайната фаза на промяна, когато вече навлизат в океана се превръщат в сребърни змиорки. Очите им се уголемяват и придобиват тъмно син цвят, променят се и пигментите на ретината им. Жълтата змиорка използва преимуществено мирисните си рецептори, докато сребърната – зрението и то специализирано за улавяне на светлина и на значителна дълбочина. Перките нарастват, кожата става дебела, страничната им линия се обособява доста добре и става видима. Настъпват и редица физиологични промени, които са свързани с промяната на химичния състав и солеността на водата, така че от придънен вид, криещ се в цепнатини и заравящ се в тинята, тя става активно плуващ, пелагичен вид. Стената на плавателния и мехур се увеличава и ефективността на газовата обмяна се увеличава многократно. Натрупват се около 20% резервни мазнини, запаси за дългото пътуване и половото узряване. Всички тези промени имат една цел – сребърната змиорка е приспособена за дълго, продължаващо хиляди километри плуване, за да може да достигне до местата за хвърляне на хайвер. По време на тази миграция змиорката спира да се храни и узрява полово, така че когато пристигне на мястото на размножаване, тя е готова да хвърли хайвера си. Змиорката притежава няколко специализирани белега, които и дават редица предимства в сравнение с другите костни риби. Тя притежава способността да издържа на хипоксия, когато в околната среда има малко съдържание на кислород, като изследвания са показали, че европейската змиорка може да живее в среда без кислород до няколко часа. Друга способност на змиорката е, че може да живее вън от водата часове, а дори и до дни, което е изключителна нейна способност сред рибите, които по принцип не могат да живеят вън от водата и се осъществява с редица физиологични особености /тънка кожа, покрит със слуз оперкулум и сърце издържащо на ацидоза/. Тези нейни способности оказват значителна помощ при миграцията и надолу по реките към океана, особено когато се натъкват на препятствия по пътя си.
Змиорката притежава няколко специализирани белега, които и дават редица предимства в сравнение с другите костни риби. Тя притежава способността да издържа на хипоксия, когато в околната среда има малко съдържание на кислород, като изследвания са показали, че европейската змиорка може да живее в среда без кислород до няколко часа. Друга способност на змиорката е, че може да живее вън от водата часове, а дори и до дни, което е изключителна нейна способност сред рибите, които по принцип не могат да живеят вън от водата и се осъществява с редица физиологични особености /тънка кожа, покрит със слуз оперкулум и сърце издържащо на ацидоза/. Тези нейни способности оказват значителна помощ при миграцията и надолу по реките към океана, особено когато се натъкват на препятствия по пътя си.

 Фотография на пресно уловени лептоцефали на японска змиорка с размер от 4 до 21 мм., показваща стадии на нарастване.
Фотография на пресно уловени лептоцефали на японска змиорка с размер от 4 до 21 мм., показваща стадии на нарастване.













